

Elektronikuljetinketjun komponentti, sekvenssi, estäjät
- 2814
- 877
- Louis Moen
Se elektronikuljetinketju Se koostuu joukosta proteiini- ja koentsyymimolekyylejä kalvon sisällä. Kuten nimestä viittaa, se vastaa elektronien kuljettamisesta NADH- tai Fadh2 -koentsyymeistä lopulliseen vastaanottimeen, joka on O2 (molekyylihappi).
Tässä kuljetusprosessissa energia, joka vapautuu siirtämällä elektroneja koentsyymeistä molekyylihappeen redox -keskuksilla, jotka ovat yhdistyneet proteiiniin, liittyy energiantuotantoon (ATP). Tämä energia saadaan protongradientin ansiosta, joka syntyy mitokondriaalisessa sisäkalvossa.
Tämä kuljetusjärjestelmä koostuu erilaisista komponenteista, joita löytyy ainakin kahdesta hapetustilasta. Jokainen niistä vähenee tehokkaasti ja reoksid elektronien liikkumisen aikana NADH: sta tai Fadh2: sta O2: een.
NAD +- ja FAD -koentsyymit vähenevät rasvahappojen hapettumisreitteissä ja sitruunahapposyklin seurauksena erilaisten substraattien hapettumisen seurauksena. Myöhemmin nämä koentsyymit hapettuu elektroniseen kuljetusketjuun.
Niin, että elektroninen kuljetusjärjestelmä koostuu toisiinsa kytkettyjen oksidien vähentämisreaktioiden sekvenssistä.
[TOC]
Ketjukomponentit
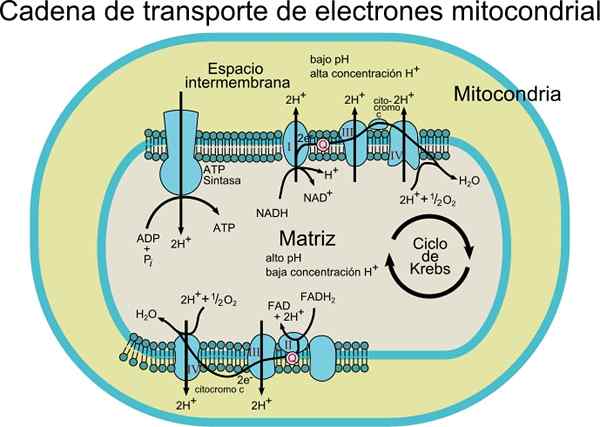
Organismin tyypistä riippuen voidaan havaita 3 - 6 komponenttia, jotka muodostavat elektronikuljetinketjun. Elektronien kuljetusprosessi ja ATP -synteesi oksidatiivisella fosforylaatiolla ovat prosesseja, jotka tapahtuvat kalvossa.
Prokaryoottisten solujen (aerobiset bakteerit) tapauksessa nämä prosessit tapahtuvat plasmamembraaniin. Eukaryot -soluissa sitä esiintyy mitokondrioiden kalvossa, joten elektronien kuljetuskomponentit löytyvät kalvon sisäosasta.
https: // giphy.com/gifs/hTksbbedquk9z7co5e
Elektronit siirretään vähitellen neljän kompleksin kautta, jotka muodostavat elektronisen kuljetusketjun.
Jokaisessa kompleksissa on useita proteesiryhmiin liittyviä proteiinikomponentteja (konjugaattiproteiinien ei -aminohappikomponentteja) redoxia, jotka mahdollistavat niiden pelkistyspotentiaalin kasvattamisen.
Lisäksi tämä kuljetusjärjestelmä koostuu erilaisista molekyylilajeista, kuten flavoproteiinista; koentsyymi, jota kutsutaan myös ubiquinonaksi (Coq tai UQ); erilaisia sytokromeja, kuten sytokromi B, C, C1, A ja A3; CU: hon kytketty proteiinit Fe-S- ja proteiiniryhmillä. Nämä molekyylit on kytketty kalvoon, lukuun ottamatta sytokromi c: tä.
Kompleksi i
https: // giphy.com/gifs/rlf1rl9gdc6356sj6w
I-kompleksi, nimeltään NADH-koentsyymi quinona oksidortaduasi tai NADH-dehydrogenaasi, koostuu noin 45 polypeptidiketjusta ja sisältää mononukleotidiflaviinimolekyylin (FMN) ja kahdeksan-yhdeksän Fe-S-ryhmän Fe-S. Kuten nimestä voi päätellä, tämä kompleksi siirtää parin elektronia NADH -koentsyymistä Coqiin.
NADH -dehydrogenaasikompleksin toiminta alkaa NADH: n liitoksella mainitun kompleksin sisäisen mitokondrioiden kalvomatriisin puolella. Sitten elektronit kuljetetaan NADH: sta FMN: ään. Seuraavaksi elektronit kulkevat pelkistetystä flavinista (FMNH2) Fe-S-proteiineihin.
FMNH2 toimii eräänlaisena siltana NADH: n ja Fe-S-proteiinien välillä, koska jälkimmäinen voi siirtää vain yhden elektronin, kun taas NADH-koentsyymi siirtää kaksi, niin että flavinit tekevät tämän yhden elektronin siirron redoxinsa ansiosta puoliksi sykynonista.
Voi palvella sinua: Monosytopoieesis: vaiheet, ominaisuudet, säätelyLopuksi, elektronit siirretään Fe-S-ryhmistä koentsyymi Q: een, joka on liikkuva elektronikuljetin, jolla on isopreneidi häntä, joka tekee siitä hydrofobisen, jonka avulla voidaan ylittää mitokondriaalisen kalvon keskusta.
Kompleksi II
![]()
II -kompleksi, joka tunnetaan paremmin nimellä sukkinaattidehydrogenaasi, on mitokondrioiden sisäisen kalvon olennainen proteiini ja se on entsyymi, joka puuttuu sitruunahapposykliin.
Tämä kompleksi koostuu kahdesta hydrofiilisestä alayksiköstä ja kahdesta hydrofobisesta HEMO B -ryhmien kanssa, jotka tarjoavat COQ.
Sitruunahapposyklissä (Krebs tai trikarboksyylihapposykli) sukkinaatti muunnetaan fumaraattiin sukkinaattidehydrogenaasin avulla, vähentäen koentsyymi -villitystä fadh2: ksi. Tästä viimeisestä koentsyymistä elektronit siirretään Fe-S-keskuksiin, jotka puolestaan siirtävät ne Coqiin.
Tämän elektroninsiirron reaktioiden aikana tavanomainen redox -potentiaali on erittäin pieni, mikä estää tarvittavan vapaan energian vapautumisen ATP: n syntetisoimiseksi.
Tämä tarkoittaa, että kompleksi II on ainoa kompleksi elektronikuljetusketjussa, joka ei kykene tarjoamaan energiaa ATP -synteesille. Tämä kompleksi on kuitenkin avainprosessissa, koska se siirtää elektroneja Fadh2: sta muuhun ketjuun.
Kompleksi III
https: // giphy.com/gifs/cjg0raiuizjds2kyu
III -kompleksi, sytokromikompleksi BC1 tai COQ -sytokromi C -reduktaasi, siirtää elektronit vähentyneestä koentsyymistä sytokromi C: hen. Tämä siirto tapahtuu yhden redox -tien kautta, joka tunnetaan nimellä Q -sykli.
Tämä kompleksi koostuu proteiinista, jolla on Fe-S ja kolme erilaista sytokromia, joissa HEMO-ryhmässä sijaitseva rautatomi vaihtelee syklisesti vähentyneiden tilojen (Fe2+) ja hapettuneen (Fe3+) välillä (Fe3+).
Sytokromit ovat elektronisen kuljetuksen hemoproteiineja, joilla on redox -aktiivisuus. Ne ovat läsnä kaikissa organismeissa, lukuun ottamatta joitain pakollisia anaerobeja.
Näissä proteiineissa on hemo -ryhmiä, jotka vuorottelevat kahden hapettumistilan välillä (Fe2+ ja Fe3+). Sytokromi C on liikkuva elektronien kuljettaja, joka liittyy heikosti mitokondrioiden sisäiseen kalvoon.
Tässä kompleksissa löydetyt sytokromit ovat sytokromit B, C ja A, 3 ovat aktiivisia redox -proteiineja, joilla on ryhmiä, joilla on erilaisia ominaisuuksia, jotka vuorottelevat niiden hapetustiloja Fe2+: n ja Fe3: n välillä+.
Sytokromi C on perifeerinen membraaniproteiini, joka toimii elektronin "sukkulana" sytokromi C1: n ja IV -kompleksin kanssa.
IV -kompleksi
Sytokromi C ja O2 ovat orgaanisen materiaalin hapettumisesta johdettujen elektronien lopullisia reseptoreita, joten IV- tai sytokromi c -oksidaasikompleksi on elektronisen kuljetusprosessin pääte -entsyymi. Tämä hyväksyy elektronit sytokromi c: stä ja siirtää ne O2: n vähentämiseen.
Kompleksin funktio on katalysoida elektronin hapettumisia, jotka ovat vähentyneen sytokromi c: n neljästä peräkkäisestä molekyylistä, ts. Samanaikaisesti vähentää O2.
Voi palvella sinua: epiteelin päällystyskudosElektronien kuljetussekvenssi
Elektronit siirretään komplekseista I ja II kompleksin III koentsyymin Q ansiosta, ja sieltä ne kulkevat IV -kompleksiin sytokromin C kautta. Kun elektronit kulkevat näiden neljän kompleksin läpi, pelkistyspotentiaali kasvaa vapauttaen energiaa, jota sitten käytetään ATP -synteesiin.
Pari elektronien siirtäminen aiheuttaa yhteensä 10 protonin siirtymistä kalvon läpi; neljä komplekseissa I ja IV ja kaksi kompleksista III.
Nadh descidrogenasa
Tämä entsyymi katalysoi NADH -koentsyymin hapettumista koentsyymin avulla. Elektronit siirtyvät NADH: sta FMN: ään, joka on kiinnittynyt kompleksin I hydrofiiliseen häntä. Fe-S-ryhmät siirtyvät elektroneihin kerrallaan. Nämä Fe-S-ryhmät vähenevät kalvoon upotettuun Coq: ksi ubikinoliin (vähentynyt Coq).
Elektronien siirron aikana CoQ: lle, neljä protonia puolestaan siirretään sisäisen kalvon läpi, väliseen tilaan. Mekanismi, jolla nämä protonit siirretään.
Elektronien siirtoprosessi tässä vaiheessa vapauttaa vapaata energiaa, erityisesti -16,6 kcal/mol.
Coq-citokromo C-reduktaasi ja Q-sykli
Koentsyymi, jonka sytokromi c hapettaa, tämän koentsyymin katalysoimassa reaktiossa. Ubikinolin (vähentynyt COQ) hapettuminen tapahtuu tietyssä kompleksin (QO tai hapettumiskohdan) tietyssä paikassa mitokondriaalikalvossa siirtämällä kaksi elektronia, joista toinen proteiiniin Fe-S-ryhmien kanssa ja toinen hemo-ryhmiin.
Q -syklissä Coq: n hapettuminen tuottaa puolikIinonia, missä elektronit siirretään Hemo B1- ja BH -ryhmiin. Vaikka tämä elektroninsiirto tapahtuu, toinen COQ hapettuu QO -alueella, toistamalla sykli.
Tämä sykli aiheuttaa kahden elektronin siirron ja vuorostaan neljän protonin siirron intermembranaaliseen tilaan vapautumisen -10,64 kcal/mol vapaata energiaa.
Citokroma C -oksidaasi
Tämä entsyymi (kompleksi IV) katalysoi sytokromi c: n (pelkistetyn) hapettumista O2: lla, joka on lopullinen elektronien vastaanottaja. Tämä siirto tuottaa H2O -molekyylin jokaiselle elektroniparille, joka on siirretty protonien translokaation lisäksi kalvon läpi.
Elektronit siirtyvät yhdestä, vähentyneestä sytokromi c: stä CUA -ionipariin, sitten ne kulkevat Hemo A -ryhmään ja lopulta saavuttavat kompleksin binukleaarikeskuksen, joka sisältää CUB- ja HEMO A3 -ionit, missä siirto neljä Elektronit esiintyvät happea.
IV -kompleksissa elementit siirtävät yksi kerrallaan elektroneja siten, että O2 vähenee asteittain, niin että joidenkin myrkyllisten yhdisteiden, kuten superoksidin, vetyperoksidin tai hydroksyyliradikaalien, vapautuminen ei tapahdu.
Tässä vaiheessa vapautuva energia vastaa -32 kcal/mol. Siirtoprosessin aikana syntynyt sähkökemiallinen gradientti ja parin elektronien aiheuttamat energiamuutokset (ΔE) neljän kompleksin läpi kulkeessa vastaa jokaisessa vaiheessa vapaata energiaa, joka on tarpeen ATP -molekyylin tuottamiseksi.
Voi palvella sinua: Membraanin taittoteoriaSuuri dehydrogenaasi
Kuten mainittiin, tällä kompleksilla on ainoa mutta tärkeä tehtävä FADH2 -elektronien käyttöönottamisesta sitruunahapposyklistä elektronien kuljetusketjuun.
Tämä entsyymi katalysoi fadh2 -koentsyymin hapettumista koentsyymi q: llä (hapettunut). Sitruunahapposyklissä, kun sukkinaatti fumaraattiin, kaksi elektronia ja kaksi protonia siirretään FAD: hen. Myöhemmin FADH2 siirtää nämä elektronit, kunnes se on COQ-kompleksin Fe-S-keskuksien kautta.
Lopuksi, COQ: sta elektronit siirretään kompleksi III: lle edellä kuvattujen vaiheiden seurauksena.
Ketjukompleksit ovat riippumattomia
Neljä kompleksia, jotka muodostavat elektronisen kuljetusketjun, ovat riippumattomia, toisin sanoen ne ovat itsenäisesti ja toimivat mitokrondriaalisessa sisäkalvossa, ja kunkin niiden liike kalvossa ei ole riippuvainen tai liitetään muihin komplekseihin.
I- ja II -kompleksit liikkuvat kalvossa siirtämällä elektronit Coq: lle, joka myös leviää kalvossa ja siirtää ne kompleksi III: lle, josta elektronit kulkevat sytokromi c: hen, joka on myös liikkuva kalvossa ja tallettaa elektroneja IV -kompleksiin.
Elektroniset kuljetusketjun estäjät
Elektronisessa kuljetusketjussa jotkut spesifiset estäjät toimivat prosessissaan. Rothenone on yleisesti käytetty hyönteismyrkky, joka on stoikiometrisesti liitetty kompleksi I: hen, välttäen Coq: n vähentämistä.
Jotkut barbituraatit tyyppiset lääkkeet, kuten pierikidiini ja amytaali, estävät kompleksia I, häiritsemällä elektronien siirtoa Fe-S-ryhmistä Coqiin.
Kompleksissa II jotkut yhdisteet, kuten illtrifluoraasetoni ja malonaatti, toimivat kilpailukykyisinä estäjinä sukkinaatissa, välttäen niiden hapettumista ja puolestaan elektronien siirtämistä villitykseen.
Jotkut antibiootit, kuten myksotiatsoli ja stigmatelliini, sitoutuvat Coq: n Q-kohtaan, estäen elektronien siirtoa koentsyymistä Q proteiinien Fe-S-keskuksiin.
Syanidi, Azida (N3-), rikkihappo ja hiilimonoksidi estävät IV-kompleksia. Nämä yhdisteet sitoutuvat hemo -ryhmiin välttäen elektronien siirtämistä kompleksi- tai happea (O2) binukleaarikeskukseen (O2).
Kun elektronikuljetinketju estetään, energiantuotanto lopetetaan oksidatiivisen fosforylaation vuoksi, aiheuttaen vakavia vaurioita ja jopa organismin kuolemaa.
Viitteet
- Alberts, b., Bray, D., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, P. (2004). Välttämätön solubiologia. New York: Garland Science. 2. painos.
- Cooper, G. M., Hausman, r. JA. & Wright, N. (2010). Solu. (PP. 397-402). Ed. Marbán.
- Devlin, t. M. (1992). Biokemian oppikirja: kliinisillä korrelaatioilla. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, c. M. (2008). Biokemia. Ed. Thomson Brooks/Cole.
- Rawn, j. D -d. (1989). Biokemia (ei. 577.1 RAW). Ed. Amerikanvälinen-McGraw-Hill
- Voet, D., & Voet, J. G. (2006). Biokemia. Ed. Pan -American Medical.
- « Sähköiset Anguila -ominaisuudet, elinympäristö, lisääntyminen
- Biozoos -ominaisuudet, morfologia, lisääntyminen, ravitsemus »