

Kadheriinien rakenne, ominaisuudet ja toiminnot
- 2870
- 774
- Shawn Stanton II
Se Kadherinat Ne ovat kalsiumista riippuvaisia glykoproteiineja, jotka ovat vastuussa solujen välisen liiton ylläpitämisestä, jotka mahdollistavat kudosten eheyden eläimiin. Kaadimaisia on yli 20 erityyppistä, kaikilla on noin 750 aminohappoa ja jotka on määritelty erityyppisistä soluista.
Kadheriinin saavuttamat soluyhdistykset ovat vakaat ajan myötä. Siksi näillä molekyyleillä on tärkeä rooli kehon muodon kehittymisessä alkion kehityksen aikana (morfogeneesi) sekä kudosten rakenteen ylläpitämisessä sekä alkion vaiheessa että aikuiselämässä.
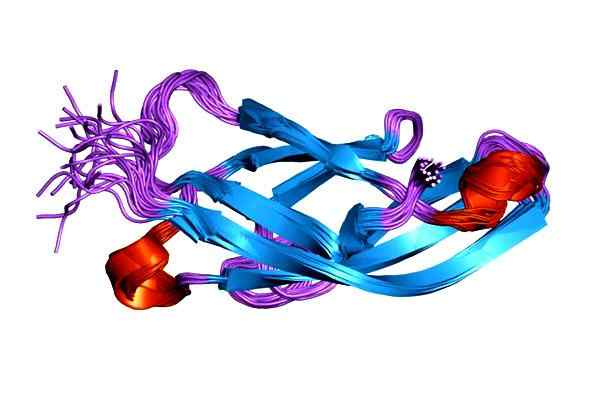
1SUH-proteiinin, E-kadherinan (epiteeli) molekyyliohjaus (epiteeli). Otettu ja toimitettu: Jawahar Swaminathan ja MSD: n henkilökunta Euroopan bioinformatiikan instituutissa [julkinen verkkotunnus]. Kadheriinin toimintahäiriö liittyy erityyppisten syövän kehitykseen. Solujen tarttumisen puute kadheriinin kautta on yksi kasvainsolujen liikkuvuuden lisääntymisen syistä.
[TOC]
Solujen tarttuvuusmolekyylit
Monisoluisissa organismeissa solujen on liityttävä osallistumaan biologisten prosessien suureen monimuotoisuuteen, jotka mahdollistavat niiden eheyden ylläpitämisen, mikä erottaa siirtomaa -yksisoluiset organismit. Näitä prosesseja ovat muun muassa hemostaasi, immuunivaste, morfogeneesi ja erilaistuminen.
Nämä molekyylit eroavat toisistaan rakenteessaan ja toiminnassaan neljässä ryhmässä: integriinit, selecinas, immunoglobuliinit ja kadherinit.
Historia
Kaadheriinin historia on hyvin lyhyt, koska he ovat tunteneet toisensa äskettäin. Siten ensimmäinen kadherina löydettiin hiirikudoksista peräisin olevista soluista vuonna 1977. Tutkijat kutsuivat tätä uvomoruliinimolekyyliä.
1980 -luvulla saavutettiin monien muiden kadheriinimolekyylien löytäminen kudoksista eri lajeista. Nämä kadherinit löydettiin kalsiumista riippuvaisista solujen aggregaatiotesteistä. Kaikki ne kuuluivat samaan molekyylien ryhmään, jota kutsutaan klassiseksi kadheriksi.
Voi palvella sinua: solujen hajoaminenViime vuosina ja molekyylibiologian edistymisen ansiosta tutkijat ovat onnistuneet tunnistamaan toisen tärkeän määrän kadheriineja, joista jotkut niiden erityistoiminnasta ei tunneta, ja sillä voi olla muita muita toimintoja kuin solujen tarttuminen.
Rakenne
Kaademat ovat glykoproteiineja, toisin sanoen molekyylejä, jotka on muodostettu proteiinin ja hiilihydraattien assosiaatiolla. Ne muodostuvat välillä 700 (yleensä 750) ja 900 aminohappoa, ja niillä on erilaiset funktionaaliset domeenit, jotka antavat sen ensin olla vuorovaikutuksessa muiden kadheriinimolekyylien kanssa ja kalsiumionien kanssa.
Funktionaaliset domeenit sallivat myös kadheriinin. Suurin osa aminohappoketjusta sijaitsee solunulkoisella alueella ja eroaa normaalisti viidessä domeenissa, nimeltään EC (EC1 - EC5).
Jokaisessa näistä domeeneista on noin 100 aminohappoa, yhden tai kaksi kalsiumyhdistyspaikkaa. Transmembraniaalinen alue on solun ulko- ja sisä- ja sisäosan välillä ja ylittää kalvon kerran.
Toisaalta kadherien osa solun sisällä on erittäin konservatiivinen ja koostuu 150 aminohapposta. Tämä domeeni sitoutuu aktiinisytoskeletoniin sytosolisten proteiinien avulla, joita kutsutaan kateniineiksi.
Kaverit
Kaadimaisia on yli 20 erityyppistä, jotka luokitellaan eri tavoin kirjoittajista riippuen. Siten esimerkiksi jotkut kirjoittajat tunnistavat kaksi ryhmää tai alaryhmää, kun taas toiset tunnustavat kuusi. Entisen mukaan kadherinit voidaan jakaa:
Klassiset kadherinit tai tyyppi I
Kutsutaan myös perinteisiksi kadheriksi. Tässä ryhmässä kadheriinit, joita kutsuttiin kankaan mukaisesti, josta ne löydettiin ensimmäistä kertaa, kuten E-kadherina (epiteeli), N-kadheriini (neuraali), P-kadheriini (istukan), L-L- Cadherina (istukan) sisältyy tähän ryhmään (istukan), L-kadherina (maksa) ja R-Cadherina (verkkokalvo). Näitä glykoproteiineja löytyy kuitenkin eri kudoksista.
Voi palvella sinua: mesosomiEsimerkiksi N-Cadherina voi sijaita myös hermostokudoksessa läsnäolon lisäksi kivesten, munuaisten, maksan ja sydämen lihaksen kudoksissa.
Epätyypilliset kadherinat tai tyyppi II
Kutsutaan myös ei -perinteiseksi tai ei -klassiseksi. Ne sisältävät demagletit ja demokoliinit, jotka muodostavat ammattiliittoja solujen välisten desmosomien tasolla. On myös protokadherinit, joille on ominaista yhteyksien puute Actinia Cytoskeletoniin.
Jotkut nämä kirjoittajat ovat erillään muista riippumattomista ryhmistä kaikki nämä kadherina erottuvat muista ei -perinteisistä ryhmistä. Muihin epätyypillisiin kadheriiniin kuuluvat T-kadherina, josta puuttuu kalvon läpäisy- ja sytoplasmiset domeenit sekä E-kadheriinin variantti, joka on solun ulkopuolella ja jota kutsutaan Evar-kadherina.
Ominaisuudet
Ne ovat kalsiumista riippuvaisia glykoproteiineja, jotka ovat melkein yksinomaan eläinkudoksissa. Suurin osa niistä on yhden vaiheen transmbrales; Toisin sanoen, että he ovat läsnä solukalvossa, ylittäen sen sivulta toiselle kerralla.
Kaademat osallistuvat pääasiassa solujen väliseen liitokseen, jolla on puoliksi haarautuneet fenotyyppiset ominaisuudet (homotyyppiset tai homofiiliset yhteydet). Näiden molekyylien (kadheriinikadheriinisidokset) saavuttamat soluvelet ovat noin 200 kertaa voimakkaampia kuin muut proteiini-proteiiniyhdistykset.
Perinteisessä kadheriinissa sytoplasminen domeeni on erittäin konservatiivinen. Tämä tarkoittaa, että sen koostumus on samanlainen eri kadheriinissa.
Funktiot
Kadheriinin päätehtävä on sallia pysyvien solujen ammattiliittojen ajan myötä, joten niillä on perustavanlaatuinen rooli prosesseissa, kuten alkion kehitys, morfogeneesi, erilaistuminen ja epiteelikudosten rakenteellinen ylläpito iholla ja suolistossa sekä aksonien muodostumisessa.
Tätä toimintoa säätelee osittain glykoproteiinin solunsisäisen domeenin terminaali -COOH. Tämä pääte on vuorovaikutuksessa molekyylien kanssa, joita kutsutaan kateniineiksi, jotka puolestaan ovat vuorovaikutuksessa solusytoskeleton elementtien kanssa.
Voi palvella sinua: polymorfonukleaariset leukosyytitKaadimojen muihin toimintoihin kuuluvat selektiivisyys (toisen soluyksikön valitseminen) ja solusignalointi, solujen polaarisuuden ja apoptoosin säätelyn muodostaminen. Jälkimmäinen on saman kehon sisäisesti solukuoleman mekanismi sen kehityksen säätelemiseksi.
Kadherinit ja syöpä
Cadherinas -toimintahäiriöt osallistuvat erityyppisten syövän kehittämiseen. Tämä toimintahäiriö voi johtua modifikaatioista kadheriinien ja kateniinien ilmentymisessä sekä solujen liiton estävien signaalien aktivoinnista.
Kun kadherien soluyhdistys epäonnistuu, tämä antaa kasvainsoluille lisätä niiden liikkuvuutta ja vapautua, ja tunkeutua sitten viereisiin kudoksiin imusolmukkeiden ja verisuonten kautta.
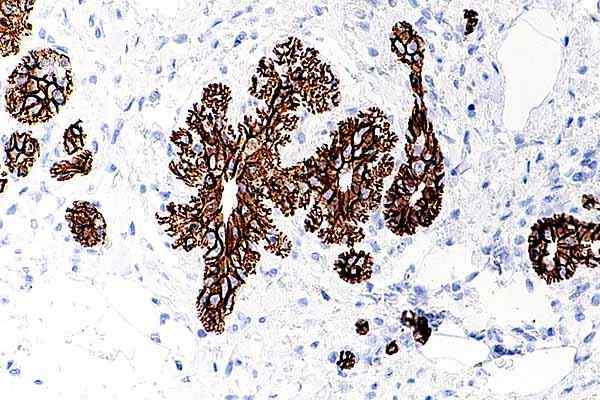 E-kadherina Benigma de Mama. Epätyypillisen lobulaarisen hyperplasian mikrokuva. Otettu ja muokattu osoitteesta: Nephron [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)].
E-kadherina Benigma de Mama. Epätyypillisen lobulaarisen hyperplasian mikrokuva. Otettu ja muokattu osoitteesta: Nephron [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)]. Kun nämä solut saavuttavat valkoiset elimet, ne tunkeutuvat ja lisääntyvät niihin, saaden invasiivisia ja metastaattisia merkkejä. Suurin osa töistä, jotka ovat yhdistäneet kadheriinin karsinogeenisiin kasvuprosesseihin, on keskittynyt E-kadheriiniin.
Tämäntyyppinen kadherina osallistuu muun muassa paksusuolen syöpään, vatsaan, rintaan, munasarjoihin ja keuhkoihin. Tämä ei kuitenkaan ole ainoa syöpään liittyvä cadherina. Esimerkiksi N-Cadherina on rooli keuhkopussin ja rabdomiosarkoomien mesotelioomissa.
Viitteet
- Kadheriini. Wikipediassa. Haettu jstk.Wikipedia.org
- D -d. Leckband & A. Prakasam (2006). Kadheriinin tarttumisen mekanismi ja dynamiikka. Vuotuinen arvostelu biolääketieteelliseen tekniikkaan.
- F. Nolletl, P. Kools P ja F. Van Roy (2000). Kadheriinin superperheen fylogeneettinen analyysi mahdollistaa kuuden suuren alaryhmän tunnistamisen useiden yksinäisten jäsenten vieressä. Journal of Molecular Biology.
- J -. Günther & e. Pedernera-Gastagian (2011). E-Cadherina: Avainpala neoplastisessa muunnoksessa. Kliininen näyttö ja tutkimuslehti.
- Lens. Petruzzelli, m. Takami & d. Humes (1999). Solujen tarttumismolekyylien rakenne ja toiminta. American Journal of Medicine.
- TAI. Cavallaro & G. Christofori (2004). Solujen tarttuminen ja kadheriinien ja IG-CAM: ien signalointi syövässä. Luontoarvostelut syöpä.
- « Iguanan ominaisuudet, lajit, elinympäristö, lisääntyminen
- Muodostumisen osteosyytit, ominaisuudet ja toiminnot »