

Kromatiinityypit, ominaisuudet, rakenne, toiminnot
- 2816
- 694
- Louis Moen
Se Kromatiini Se on DNA: n ja proteiinien muodostama kompleksi, ainutlaatuinen eukaryoottisissa organismeissa. Suhteen suhteen se sisältää melkein kaksinkertaisen proteiinin geneettisestä materiaalista. Tämän kompleksin tärkeimmät proteiinit ovat histonit - pienet proteiinit, joilla on positiivinen kuormitus, jotka sitoutuvat DNA: han sähköstaattisella vuorovaikutuksella. Lisäksi kromatiinilla on yli tuhat proteiinia erilainen kuin histonit.
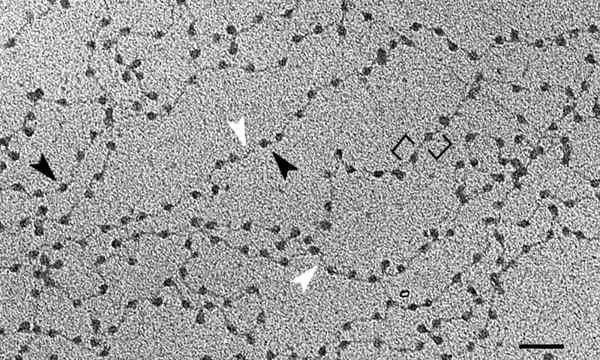
Kromatiinin perusyksikkö on nukleosomi, joka koostuu histonien ja DNA: n liitosta. Tämä ordinaatio muistuttaa kaulakorun tilejä. Saatuaan läpi kaikki korkeammat organisaatiotasot, saavutamme kromosomit.
Lähde: kromatiini_nucleofilaments.PNG: Chris Woodcockderivepative Work: GoutTegd [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Kromatiinin rakenne liittyy läheisesti geeniekspression hallintaan. Päätyyppejä on kahta: euchromatiini ja heterokromatiini.
Eukromatiinille on ominaista, että sillä on pieni tiivistysaste, mikä tarkoittaa suurta transkriptiotasoa. Sitä vastoin heterokromatiini on transkriptionaalisesti inaktiivinen, johtuen niin korkeasta tiivistysasteesta, että se esittelee.
Rakenteellisesti on tiettyjä epigeneettisiä merkkejä molempien kromatiinien histoneista. Vaikka euchromatiini liittyy asetylointiin, heterokromatiini liittyy kemiallisten ryhmien laskuun.
On joitain heterokromatiinialueita, joilla on rakennefunktioita, kuten telomeerit ja sentromeerit.
[TOC]
Historiallinen näkökulma
Geneettisen materiaalin ja sen rakenteellisen organisaation tutkimus alkaa vuonna 1831, kun tutkija Robert Brown kuvaa ydintä. Yksi tämän löytön välittömistä kysymyksistä oli tutkia rakenteen biologista ja kemiallista luonnetta.
Nämä kysymykset alkoivat selvittää vuosina 1870–1900 Friedrich Miescherin kokeilla, jotka esittelevät sanan nukleiini. Walther Flemming kuitenkin muuttaa termiä ja käyttää Kromatiini Viitata ydinaineen.
Ajan myötä aloitat syvemmän tiedon geneettisestä materiaalista ja sen ominaisuuksista. Vasta vuonna 1908, kun Italia Pasquale Baccarinin tutkija toteaa, että kromatiini ei ole homogeeninen ja onnistuu visualisoimaan pieniä kappaleita ytimen sisällä.
Emil Heitz ehdotti alun perin kromatiinien - euchromatiinin ja heterokromatiinin - tyypit vuonna 1928. Tämän luokituksen perustamiseksi Heitz perustui värjäyksen käyttöön.
Vuonna 1974 biologi Roger Kornberg ehdotti mallia geneettisen materiaalin järjestämiselle nukleosomeina tunnetuissa rakenteissa, hypoteesi, joka on empiirisesti vahvistettu Markus Noll -kokeilla.
Mikä on kromatiini?
Kromatiinikomponentit: DNA ja proteiinit
Kromatiini on nukleoproteiini, joka muodostuu geneettisen materiaalin yhdistämisestä - heterogeenisen proteiinisarjan kanssa. Tämä assosiaatio on erittäin dynaaminen ja hankkii monimutkaisen kolmiulotteisen konformaation, jonka avulla se voi suorittaa säätely- ja rakennefunktiot.
Voi palvella sinua: ProfaseYksi tärkeimmistä kromatiiniproteiineista on histonia, jotka ovat melkein yhtä suuret kuin DNA.
Histonit ovat emäksisiä proteiineja, jotka ovat huomattavasti säilyneitä orgaanisten olentojen evoluutiohistoriassa - ts. Histonimme eivät vaihtele paljon verrattuna muiden nisäkkäiden, jopa toisen kaukaisempien eläinten fylogeneettisesti.
Histonien taakka on positiivinen, joten ne voivat olla vuorovaikutuksessa sähköstaattisten voimien läpi DNA: ssa olevan fosfaattirurungin negatiivisen kuormituksen kanssa. Histoneja on viisi, nimittäin: H1, H2A, H2B, H3 ja H4.
On myös sarja proteiineja, jotka ovat erilaisia kuin DNA -tiivistykseen osallistuvat histonit.
DNA -tiivistys: Nukleosomit
Kromatiinin emäksinen yhtenäisyys on nukleosomit - toistuvat rakenteet, jotka on muodostettu DNA: sta ja histoneista, konformaatiosta, jonka löydämme koko geneettisestä materiaalista.
Kaksinkertainen DNA -potkuri on valssattu kahdeksassa histonikompleksissa, joka tunnetaan nimellä Histonasokki. Molekyyli rullataan suunnilleen kahdessa kierroksessa, jota seuraa lyhyt alue (välillä 20 - 60 emäsparia), joka erottaa nukleosomit toisistaan.
Tämän organisaation ymmärtämiseksi meidän on otettava huomioon, että DNA -molekyyli on erittäin pitkä (noin 2 metriä) ja että se on ilmoittautunut järjestämällä asettua ytimeen (jonka halkaisija on 3–10 µm). Lisäksi sen on oltava käytettävissä replikointia ja transkriptiota varten.
Tämä tavoite saavutetaan DNA: n eri tiivistystasoilla, koska se on ensimmäinen heistä edellä mainitut nukleosomit. Nämä muistuttavat helmen kaulakorun tilejä. Noin 150 DNA -pohjaparia rullataan kotona "tili".
Bakteereissa ei ole todellisia tarinoita. Sitä vastoin on sarja proteiineja, jotka muistuttavat histoneja, ja oletetaan, että ne edistävät bakteerien DNA -pakkauksia.
Korkeampi organisaatio
Kromatiinin organisointia ei ole rajoitettu nukleosomien tasolla. Tämä proteiini- ja DNA -assosiaatio on ryhmitelty paksumpaan rakenteeseen, joka on noin 30 nm - tästä paksuudesta johtuen sitä kutsutaan ”30 nm: n kuitutasoksi”.
30 nm: n paksuuteen järjestetty kromatiini järjestetään vuorotellen silmukoiden muodossa, jotka ulottuvat eräänlaisena proteiinin luonto -telineessä (ei histonit).
Tämä on malli, jota käsitellään tällä hetkellä, vaikka monimutkaisempien tiivistysmekanismien olemassaoloa voidaan odottaa. Viimeinen organisaatio koostuu kromosomista.
Se voi palvella sinua: Cilia: Ominaisuudet, rakenne, toiminnot ja esimerkitVirheet kromatiinin organisaatiossa
Geneettisen materiaalin tiivistäminen ja organisointi on välttämätöntä useille biologisille toiminnoille. Erilaisia sairauksia on liitetty virheisiin kromatiinin rakenteessa, mukaan lukien alfa-talaasia, joka on kytketty X-kromosomiin, Rubinstein-Taybi-oireyhtymään, arkku-alhaiseen oireyhtymään, Rett-oireyhtymään, muun muassa.
Kromatiinityypit

Solussa on kahta kromatiinityyppiä, jotka paljastetaan värjäyksen levittämisellä: euchromatiinia ("tosi" kromatiinia) ja heterokromatiinia. Ensimmäisessä tapauksessa värjäys havaitaan heikosti, kun taas toisessa värjäys on voimakas.
Tämä DNA: n rakenteellinen organisaatio on ainutlaatuinen eukaryoottisille organismeille ja on ratkaisevan tärkeä kromosomien käyttäytymiselle ja geeniekspression säätelemiselle.
Jos arvioimme molempien kromatiinien osuuksia solussa, joka on rajapinnassa, havaitsemme, että noin 90% kromatiinista on euchromatiinia ja loput 10% vastaa heterokromatiinia. Seuraavaksi kuvaamme jokaista tyyppiä yksityiskohtaisesti:
Yllyttää. Heterokromatiini
Ominaisuudet
Tärkein ero molempien kromatiinityyppien välillä liittyy molekyylin tiivistymisasteeseen tai "pakkaamiseen" tietyissä vaiheissa solun jakautumisessa.
Vaikka rajapinnassa geneettinen materiaali näyttää olevan satunnaisesti dispergoitunut, se ei ole tällä tavalla.
Tässä vaiheessa on merkittävä organisaatio, jossa voit nähdä differentiaalisen osion kromosomaalisesta materiaalista ytimen sisällä.
Kromosomien DNA ei ole kietoutunut muiden kromosomien DNA -juosteen kanssa, ja jäännökset ovat spesifisiä alueita, joita kutsutaan kromosomaalisiksi alueiksi. Tämä organisaatio näyttää edistävän geeniekspressiota.
Heterokromatiini on voimakkaasti tiivistynyt, koska se on vähän pääsyä transkriptiokoneille - joten niitä ei kirjoiteta. Lisäksi se on huono sen esittämien geenien määrän suhteen.
Heterokromatiinin tyypit
Tietyt heterokromatiinialueet ovat pysyviä kaikissa solulinjoissa - eli, aina Se käyttäytyy kuin heterokromatiini. Tämän tyyppistä heterokromatiinia kutsutaan konstitutiiviseksi. Esimerkki tästä ovat kromosomien tiivistetyt alueet, joita kutsutaan sentromeeriksi ja telomeereiksi.
Sitä vastoin on heterokromatiinin osia, jotka voivat vaihdella tiivistystasoa vasteena kehitysmallien tai ympäristömuuttujien muutoksille.
Uusien tutkimusten ansiosta tämä visio muotoillaan uudelleen ja nyt on todisteita siitä, että konstitutiivinen heterokromatiini on myös dynaaminen ja kykenevä reagoimaan ärsykkeisiin.
Rakenne
Yksi tekijöistä, jotka määrittelevät kromatiinin rakenteen, ovat histonien kemialliset modifikaatiot. Kromatiinin tapauksessa, joka on transkriptionaalisesti passiivinen, heillä on hypoasetoituja histoneja.
Voi palvella sinua: peroksidaasit: rakenne, toiminnot ja tyypitAsetyyliryhmien määrän väheneminen liittyy geenien hiljaisuuteen, koska lisinien positiivinen kuormitus ei masskaa, mikä mahdollistaa vahvan sähköstaattisen vuorovaikutuksen DNA: n ja histonien välillä.
Toinen epigeneettinen tuotemerkki on metylaatio. Koska metyyliryhmän lisääminen ei kuitenkaan modifioida proteiinin kuormaa, sen seuraus (aktivoi tai deaktivointi geenit) ei ole niin ilmeinen ja riippuu histonin alueesta, jossa tuotemerkki sijaitsee.
Empiirisesti on havaittu, että H3K4me3: n ja H3K36me3: n metylaatio liittyy geeniaktivaatioon ja H3K9me3: n ja H3K27me3: n aktivaatioon.
Funktiot
Konstitutiivisen heterokromatiinin esimerkissä mainitsemme sentromeerin. Tällä kromosomaalisella alueella on rakenteellinen rooli ja se myötävaikuttaa kromosomien liikkumiseen sekä mitoottisten että meioottisten solujen jakautumisen tapahtumien aikana.
II. Euchromatiini
Ominaisuudet
Vastoin heterokromatiinia, euchromatiini on vähemmän kompakti molekyyli, joten transkriptiokoneella on helppo pääsy (erityisesti entsyymi -RNA -polymeraasi) ja se voidaan ekspressoida aktiivisesti geneettisillä reiteillä.
Rakenne
Aktiivisen kromatiinin nukleosomin rakenteelle transkription näkökulmasta on tunnusomaista asetyloidut histonit. Monometyyli -liisiinien läsnäolo liittyy myös geenin aktivaatioon.
Asetyyliryhmän lisääminen tähän lysiinijätteeseen histonista neutraloi mainitun aminohapon positiivisen kuorman. Tämän muutoksen välitön seuraus on histonin ja DNA: n sähköstaattisten vuorovaikutusten vähentäminen, mikä tuottaa laisemman kromatiinin.
Tämä rakenteellinen modifikaatio mahdollistaa geneettisen materiaalin vuorovaikutukset transkriptionaalisten koneiden kanssa, jolle on ominaista erityisen tilaa vievä.
Funktiot
Eukromatiini kattaa kaikki aktiiviset geeni. Siksi toiminnot ovat yhtä laajoja kuin kyseisten geenien toiminnot.
Viitteet
- Grewal, s. Yllyttää., & Moozed, D. (2003). Geeniekspression heterokromatiini ja epigeneettinen hallinta. Tiede, 301(5634), 798-802.
- Jost, k. Lens., Bertulat, b., & Cardoso, m. C. (2012). Heterokromatiini ja geenin sijainti: Sisällä, ulkopuolella, mikä tahansa puoli?. Kromosomi, 121(6), 555-563.
- Lewin, b. (2008). IX -geenit. Jones ja Bartlett Publishers.
- Tollefsbol, t. JOMPIKUMPI. (2011). Epigenetiikan käsikirja. Akateeminen lehdistö.
- Wang, J., Jia, S. T., & Jia, S. (2016). Uudet näkemykset heterokromatiinin säätelystä. Genetiikan suuntaukset: TIG, 32(5), 284-294.
- Zhang, P., Torres, k., Liu, x., Liu, c. G., & Pollock, R. JA. (2016). Katsaus kromatiinia säätelevistä proteiineista soluissa. Nykyinen proteiini- ja peptiditiede, 17(5), 401-410.
- « Ergastiset aineet luokittelu ja biologinen merkitys
- Yucca Filifera -ominaisuudet, taksonomia, elinympäristö, käyttö »