

Sinkkisormet, rakenne, luokittelu, toiminto
- 1084
- 179
- Arthur Terry II
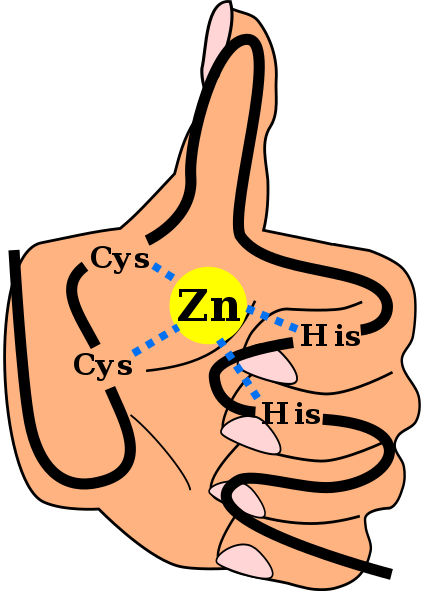
Sinkkisormen domeenin C2H2 ensisijainen rakenne, mukaan lukien sinkki -ionin koordinoivat linkit ja "käden ja sormen" taustalla. Angel Herraez, Wikimedia Commons Se Sinkkisormit (Zf) ovat rakenteellisia aiheita, jotka ovat läsnä suuressa määrässä eukaryooteja. He kuuluvat metalloproteiiniryhmään, koska he kykenevät yhdistämään sinkkimetalli -ionin, jota he vaativat sen toimintaan. Ennustetaan, että yli 1500 ZF -domeenia esiintyy noin 1000 erilaisessa proteiinissa ihmisillä.
Termi sinkkisormi tai ”sinkkisormi” keksivät ensimmäisen kerran vuonna 1985 Miller, McLachlan ja Klug tutkiessaan yksityiskohtaisesti pieniä domeeneja transkription tekijän TFIIIA DNA: n DNA: n pieniä domeeneja Xenopus laevis, muut kirjoittajat ovat kuvanneet muutama vuosi ennen.
Proteiinit, joilla on ZF -motiivit toiset.
Rakenne
ZF -aiheiden rakenne on erittäin säilynyt. Yleensä näillä toistuvilla alueilla on 30 - 60 aminohappoa, joiden sekundaarinen rakenne on kuin kaksi beeta -antiparallel -arkkia, jotka muodostavat haarukan ja alfa -välityspalvelimen, joka on merkitty nimellä ββα.
Tämä sekundaarinen rakenne stabiloituu hydrofobisilla vuorovaikutuksilla ja kahden kysteiinin ja kahden histidiinijätteen antaman sinkkiatomin koordinaatiolla (CYS2Hänen2-A. On kuitenkin ZF, joka voi koordinoida useampaa kuin yhtä sinkkiä atomia ja muita, joissa CYS: n ja hänen jätteidensä järjestys vaihtelee.
Voi palvella sinua: palmitholiisiinihappo: rakenne, toiminnot, missä se onZFS voidaan toistaa erässä, lineaarisesti konfiguroitu samassa proteiinissa. Kaikilla on samanlaiset rakenteet, mutta ne voivat erottaa itsensä kemiallisesti avainten aminohappojätteiden muunnelmat heidän toimintojensa toteuttamiseksi.
Yleinen ominaisuus ZF: n välillä on sen kyky tunnistaa DNA- tai RNA -molekyylit, joilla on eri pituus, minkä vuoksi niitä pidettiin alun perin vain transkriptiivisina tekijöinä.
Yleensä tunnistus on 3PB: n alueita DNA: ssa ja saavutetaan, kun ZF -domeenin proteiini esittelee alfapotkurin DNA -molekyylin pääuraan.
Luokittelu
On olemassa erilaisia ZF -syitä, jotka eroavat toisistaan luonteeltaan ja erilaiset avaruuskokoonpanot, jotka saavuttavat koordinaatioyhteydet sinkkiatomiin. Yksi luokituksista on seuraava:
C2H2
Tämä on syy ZF: stä yleisesti. Useimmat syyt c2H2 Ne ovat spesifisiä vuorovaikutukseen DNA: n ja RNA: n kanssa, mutta niiden on havaittu osallistuvan proteiini-proteiini-vuorovaikutuksiin. Heillä on 25–30 aminohappojätettä ja he ovat nisäkkäiden suurimman säätelyproteiinien perheessä.
C2H
Vuorovaikutuksessa RNA: n ja joidenkin muiden proteiinien kanssa. Niitä havaitaan pääasiassa osana joitain retroviruksen kapselin proteiineja, jotka tekevät yhteistyötä virus -RNA: n pakkauksessa heti replikaation jälkeen.
C4 (silmukka tai nauha)
Tällä motiivilla varustetut proteiinit ovat entsyymejä, jotka vastaavat DNA: n replikaatiosta ja transkriptiosta. Hyvä esimerkki näistä voi olla faagien ja T7: n raa'at entsyymit.
C4 (Gata -perhe)
Tämä ZF -perhe sisältää transkriptiotekijöitä, jotka säätelevät tärkeiden geenien ilmentymistä lukuisissa kudoksissa solujen kehityksen aikana. Esimerkiksi gata-2 ja 3 tekijää ovat mukana hematopoieesissa.
Se voi palvella sinua: ATP (adenosiinitriffosfaatti)C6
Nämä domeenit ovat tyypillisiä hiivoille, erityisesti Gal4 -proteiinille, joka aktivoi geenien ja melibious -käyttämiseen osallistuvien geenien transkription.
Sinkkisormet (c3HC4-C3H2C3-A
Näissä erityisissä rakenteissa on 2 ZF -domeenin alatyyppiä (C3HC4 ja c3H2C3) ja ovat läsnä lukuisissa eläin- ja kasviproteiineissa.
Niitä löytyy proteiineista, kuten Rad5, jotka osallistuvat DNA: n korjaamiseen eukaryoottisissa organismeissa. Niitä löytyy myös Rag1: stä, välttämättömiä immunoglobuliinien uudelleenkonfigurointiin.
H2C2
Tämä ZF -domeeni on erittäin säilynyt retrovirus- ja retrotransposon -integraaleissa; Kun liityt valkoista proteiinia, se aiheuttaa konformaatiomuutosta samassa.
Funktiot
ZF -domeeniproteiinit palvelevat erilaisia tarkoituksia: niitä löytyy ribosomaalisista proteiineista tai transkriptiivisista adaptereista. Ne on havaittu myös olennaisena osana hiivan RNA -polymeraasi II rakennetta.
Ne näyttävät olevan mukana solunsisäisessä sinkin homeostaasissa ja apoptoosin tai ohjelmoidun solukuoleman säätelyssä. Lisäksi on joitain ZF -proteiineja, jotka toimivat chaperoneina muiden proteiinien taittamiseen tai kuljettamiseen.
Lipidiliitot ja perustavanlaatuinen rooli proteiini-proteiini-vuorovaikutuksissa ovat myös ZF-domeenien erinomaisia funktioita joissain proteiineissa.
Bioteknologinen merkitys
Vuosien mittaan ZF -domeenien rakenteellinen ja toiminnallinen ymmärtäminen on mahdollistanut suuren tieteellisen edistyksen, joka merkitsee niiden ominaisuuksien käyttöä bioteknologisiin tarkoituksiin.
Koska joillakin ZF -proteiineilla on suuri spesifisyys tietyistä DNA -domeeneista, tällä hetkellä on sijoitettu paljon ponnisteluja erityiseen ZF -suunnitteluun, joka voi tarjota arvokasta edistystä ihmishoidossa ihmisillä.
Se voi palvella sinua: Kiinan kasvisto ja eläimistö: edustavat lajitMielenkiintoisia bioteknologisia sovelluksia syntyy myös proteiinisuunnittelusta, jonka ZF on muokattu geenitekniikan avulla. Halutusta päästä riippuen, osa näistä voidaan muokata lisäämällä sormet "Poli sinkki" -sormilla, jotka kykenevät käytännössä tunnistamaan kaikki DNA -sekvenssit suurella affiniteettilla ja spesifisyydellä.
Genominen painos modifioituilla nukleilla on yksi lupaavimmista sovelluksista. Tämäntyyppinen painos tarjoaa mahdollisuuden suorittaa geneettisen toiminnan tutkimuksia suoraan kiinnostavassa järjestelmässä.
Geenitekniikka, joka käyttää modifioituja ZF -nukleasoja. Näitä nukloja on käytetty endogeenisen geenin korjaamiseen, joka tuottaa Herbisidal -resistenttejä muotoja tupakkakasveissa.
Nukloja, joilla on ZF, on käytetty myös geenien lisäämiseen nisäkkäiden soluihin. Kyseisiä proteiineja käytettiin generoimaan joukko isogeenisiä hiirisoluja, joissa oli sarja alleeleja, jotka on määritelty endogeeniselle geenille.
Tällaisella prosessilla on suora sovellus merkinnöissä ja uusien alleelisten muotojen luomisessa rakenteen ja funktiosuhteiden tutkimiseksi natiivissa ekspressio -olosuhteissa ja isogeenisissä ympäristöissä.
Viitteet
- Berg, J. M. (1990). Sinkkisormen alueet: hypoteesit ja nykyinen tieto. Biofysiikan ja biofysikaalisen kemian vuosikatsaus, 19(39), 405-421.
- Kluska, k., Adamczyk, J., & Krȩzel, a. (2017). Sinkkisormien metallin sitoutumisominaisuudet luonnollisesti altenmetallin sitoutumiskohdalla. Metallomia, 10(2), 248-263.