

Solunjakotyypit, prosessit ja merkitys
- 1187
- 133
- Gustavo Runte DVM
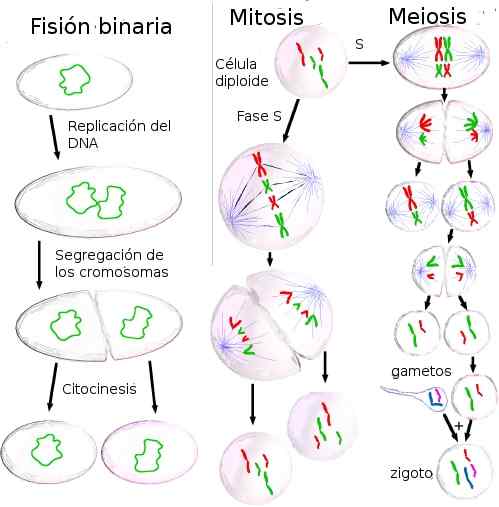
Se solunjako Prosessi antaa kaikkien elävien organismien kasvaa ja lisääntyä. Prokaryooteissa ja eukaryooteissa solunjakojen tulos ovat tytärsolut, joilla on sama geneettinen tieto kuin alkuperäisellä solulla. Tämä tapahtuu, koska ennen jakoa DNA: n sisältämät tiedot kaksinkertaistuvat.
Prokaryooteissa divisioona tapahtuu binaarisella fissiolla. Useimpien prokaryootien genomi on pyöreä DNA -molekyyli. Vaikka näillä organismeilla ei ole ydintä, DNA on tiivistetyssä muodossa, jota kutsutaan nukleoidiksi, joka eroaa sitä ympäröivästä sytoplasmasta.
Lähde: Rtama [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]] Eukaryooteissa jako tapahtuu mitoosin ja meioosin kautta. Eukaryoottinen genomi koostuu suurista määristä organisoitua DNA: ta ytimessä. Tämä organisaatio perustuu DNA -pakkauksiin, joissa on proteiineja, muodostaen kromosomeja, jotka sisältävät satoja tai tuhansia geenejä.
Hyvin monimuotoisilla eukaryooteilla, sekä yksisoluisilla että metatsoareilla, on elinkaaria, jotka vuorottelevat mitoosi ja meioosi. Nämä syklit ovat niitä, joilla on: a) gammainen meioosi (eläimet, jotkut sienet ja levät), b) sygoottinen meioosi (jotkut sienet ja alkueläimet); ja c) Vaihto pelaamisen ja sygoottisen meioosin (kasvien) välillä.
[TOC]
Kaverit
Solunjako voi tapahtua binaarisella fissiolla, mitoosilla tai meioosilla. Seuraava kuvaa jokaista tämän tyyppisiin solujen jakautumiseen liittyviä prosesseja.
Binaarinen fissio
 Prokarioottinen fissio, binaarinen fissio, on aseksuaalinen lisääntymismuoto.
Prokarioottinen fissio, binaarinen fissio, on aseksuaalinen lisääntymismuoto. Binaarinen fissio koostuu solun jakautumisesta, joka aiheuttaa kaksi tytärsolua, jokaisella on identtinen kopio alkuperäisestä solu -DNA: sta.
Ennen prokaryoottisen solun jakautumista DNA: n replikaatio tapahtuu, joka alkaa kaksoisketjun DNA: n tietyssä paikassa, jota kutsutaan replikaation alkuperästä. Replikaatioentsyymit liikkuvat kohti molempiin lähtökohtia, tuottaen kopion jokaisesta kaksiketjusta DNA -ketjuista.
DNA: n replikaation jälkeen solu pidentyy ja DNA erotetaan soluun. Välittömästi uusi plasmamembraani alkaa kasvaa solun keskellä muodostaen väliseinän.
Tätä prosessia helpottaa FTSZ -proteiini, joka on evoluutio erittäin säilynyt prokaryooteissa, mukaan lukien Archaea. Lopuksi solu on jaettu.
Solusykli ja mitoosi
Vaiheet, joiden kautta kahdesta peräkkäisestä solujen jakautumisesta on eukaryoottinen solu, tunnetaan solusyklinä. Solusyklin kesto vaihtelee muutamasta minuutista kuukausiin solutyypistä riippuen.
Solusykli on jaettu kahteen vaiheeseen, nimittäin vaihe M ja rajapinta. Kaksi prosessia, joita kutsutaan mitoosiksi ja sytokiineille, esiintyy M -vaiheessa. Mitoosi koostuu ydinosastosta. Alkuperäisessä ytimessä läsnä olevat sama määrä ja tyypit löytyvät lasten ytimistä. Monisoluisten organismien somaattiset solut on jaettu mitoosilla.
Sytokiinit koostuu sytoplasman jakautumisesta tytärsolujen muodostamiseksi.
Rajapinnassa on kolme vaihetta: 1) G1, solut kasvavat ja viettävät suurimman osan ajastaan tässä vaiheessa; 2) s, genomin päällekkäisyys; ja 3) G2, mitokondrioiden ja muiden organelien replikaatio, kromosomien kondensaatio ja mikrotubulusten kokoonpano, muun muassa.
Mitoosivaiheet
Mitoosi alkaa G2 -faasin lopussa, ja se on jaettu viiteen vaiheeseen: profasi, lupaus, metafaasi, anafaasi ja tefelaasi. Ne kaikki tapahtuvat jatkuvasti.
Voi palvella sinua: Solun viestintä: Tyypit, merkitys, esimerkkejäProfaatti
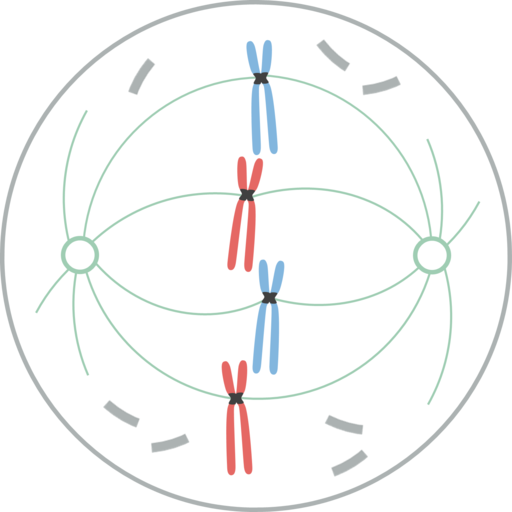 Profaatti. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons
Profaatti. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons Tässä vaiheessa mitoottisen karan tai mitoottisen laitteen kokoonpano on päätapahtuma. Profasi alkaa kromatiinin tiivistyksellä, muodostaen kromosomit.
Jokaisessa kromosomissa on pari sisarekromaatit, identtisen DNA: n kanssa, jotka ovat läheisesti yhteydessä niiden sentromeerien naapurustoon. Proteiinikompleksit, joita kutsutaan kohesiiniksi, osallistuvat tähän liittoon.
Jokainen sentromeeri on kytketty Cinetocoroon, joka on proteiinikompleksi, joka sitoutuu mikrotutkiin. Nämä mikrotubulot sallivat jokaisen kromosomien kopion määrittämisen tytärsoluille. Mikrotubulukset radian solun molemmista päistä ja muodostavat mitoottisen laitteen.
Eläinsoluissa, ennen profaasia, keskittymisen päällekkäisyys tapahtuu, mikä on mikrotubulusten tärkein järjestämiskeskus ja paikka, jossa vanhempi ja pojan keskipisteet sijaitsevat. Jokainen keskitys saavuttaa solun vastakkaisen navan, mikä muodostaa mikrotubulos -sillan niiden väliin nimeltään mitoottinen laite.
Viimeisimmissä evoluutiokasveissa, toisin kuin eläinsolut, keskuksia ei ole ja mikrotubulusten alkuperä ei ole selvää. Vanhempien evoluutioperusteiden fotosynteettisissä soluissa, kuten vihreät levät, on keskuksia.
Lupaus
Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Mitoosin tulisi taata kromosomien segregaatio ja ydinhuokos- ja nukleolikompleksin ydinkuoren jakautuminen. Riippuen siitä, katoaako ydinkääre (in) vai ei, ja IN: n denstegratiivisuusaste, mitoosi vaihtelee suljettuun kokonaan avoimeen.
Esimerkiksi S. Cerevisae Mitoosi on suljettu, vuonna -Lla. Nidulans Se on puoliksi avoin, ja ihmisillä se on auki.
Suljetussa myitoosissa karan napakappaleet ovat ydinkuoren sisällä, jotka muodostavat ydin- ja sytoplasmisten mikrotubulusten ytimet kohdat. Sytoplasmiset mikrotubulukset ovat vuorovaikutuksessa solun aivokuoren kanssa ja kromosomien kanssa.
Puoli -abserti -myitoosissa, koska IN on osittain desensamblada, ydintilaa tunkeutuu keskuksista nukkeutuneiden mikrotubulusten ja kahden aukon kautta, muodostuminen.
Avoa mitoosissa täydellinen de -salamblate esiintyy, mitoottinen laite on valmis ja kromosomit alkavat siirtyä solun keskustaa kohti.
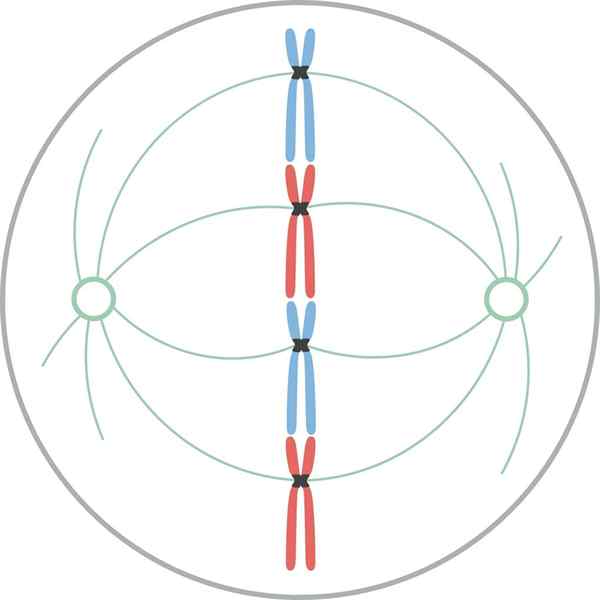
Metafaasi
Kromosomit kohdistuvat solun päiväntasaajan levyyn myyttisen metafaasin aikana Metafaasissa kromosomit ovat linjassa solun soluun. Kuvitteellinen taso, joka on kohtisuorassa karan akseliin, joka kulkee solun sisätilan läpi, kutsutaan metafaasiruokaksi.
Nisäkässoluissa mitoottinen laite on järjestetty keskimmäiseen mitoottiseen karaan ja pariin Osteres. Mitoottinen kara koostuu symmetrisestä kahdenvälisestä mikrotubulusten kimppusta, joka on jaettu solun soluun, muodostaen kaksi vastakkaista puolikkaata. Ásteres koostuu mikrotubulusten ryhmästä jokaisessa karan napassa.
Mitoottisessa laitteessa on kolme mikrotubulusryhmää: 1) astraali, jotka muodostavat rsterin, alkavat keskittämisestä ja säteilevät kohti solukorteksia; 2) Cinetocorosta, jotka sitoutuvat kromosomeihin Cinetocoron läpi; ja 3) polaarinen, joka interdignoi vastakkaisen navan mikrotubuluksia.
Kaikissa yllä kuvattuissa mikrotubuluksissa päät (-) ovat suunnattuja keskuksiin.
Karara on kasvisoluissa, jos keskitytään, kara on samanlainen kuin eläinsolujen. Kara koostuu kahdesta puolikkaasta, joilla on vastakkainen napaisuus. Päät (+) löytyvät päiväntasaajan levystä.
Se voi palvella sinua: podosyytit: ominaisuudet, histologia ja toiminnotAnafasi
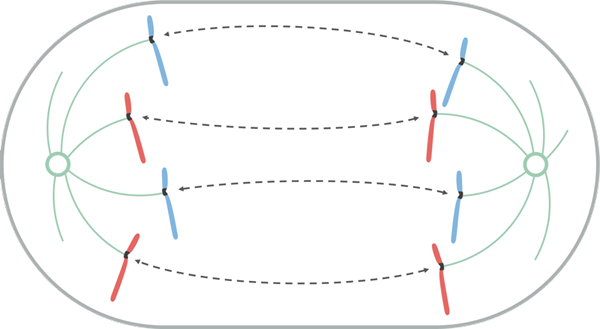 Lähde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons
Lähde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons Anafaasi on jaettu aikaisin ja myöhään. Varhaisessa anafaasissa sisarkromatidien erottaminen tapahtuu.
Tämä erotus tapahtuu, koska unionin ylläpitävät proteiinit ovat pukkia ja koska Cynetocoro -mikrotubulukset lyhenevät. Kun sisarkromatidien pari erotetaan, niitä kutsutaan kromosomeiksi.
Kromosomien siirtymisen aikana napoihin kohti Cinetocoro liikkuu saman cynetokoron mikrotubuluksia pitkin kuin sen pää (+) dissosiaatiot. Tämän vuoksi kromosomien liikkuminen mitoosin aikana on passiivinen prosessi, joka ei tarvitse motorisia proteiineja.
Myöhäisessä anafaasissa pylväiden suurempi erotus tapahtuu. KRP -proteiini, joka on kytketty polaaristen mikrotubulusten äärimmäiseen (+), saman päällekkäisyyden alueella, marssii viereisen polaarisen mikrotubulus -antiparallelelin päätä (+). Siten KRP työntää viereisen polaarisen mikrotubuluksen päätä kohti (-).
Kasvisoluissa, kromosomin erottamisen jälkeen. Tämä rakenne mahdollistaa sytotsineettisen laitteen alkamisen, nimeltään Framoplasto.
Telfaasi
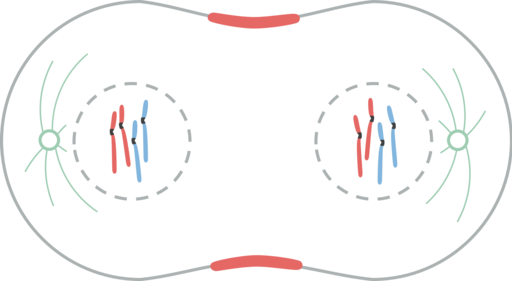 Telfaasi. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)]
Telfaasi. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Telophasessa tapahtuu useita tapahtumia. Kromosomit saavuttavat pylväät. Cinetocoro katoaa. Polaariset mikrotubulukset jatkavat itseään pidentämällä, solun valmistelua sytokiinille. Ydinkuori muodostuu jälleen äidin käärettävistä fragmenteista. Nukleoli ilmestyy uudelleen. Kromosomit ymmärretään väärin.
Sytokiinia

Civotsinesis on solusyklin vaihe, jonka aikana solu on jaettu. Eläinsoluissa sytosiinit esiintyy aktiinifilamentin supistumishihnalla. Nämä filamentit liukuvat toistensa yli, hihnan halkaisija vähenee ja klivajen ura muodostuu solun ympärysmitasta.
Koska supistuminen jatkuu, ura syvenee ja muodostuu solujen välinen silta, joka sisältää keskimääräisen rungon. Solunvälisen sillan keskialueella ovat mikrotubulusten palkit, jotka peittävät elektrodense -matriisin.
Solunvälisen sillan repeämä post-mitoottisten sisarisolujen välillä tapahtuu absission kautta. Alleja on kolme tyyppiä: 1) mekaaninen repeämämekanismi; 2) arkistomekanismi sisäisillä vesikkeleillä; 3) Fission plasmamembraanin supistuminen.
Kasvisoluissa membraanikomponentit kootaan sisälle ja solun plakki muodostuu. Tämä levy kasvaa plasmamembraanin pinnalle, sulautuen sen kanssa ja jakamalla solun kahteen. Sitten selluloosa kerrostuu uuteen plasmamembraaniin ja muodostaa uuden soluseinämän.
Meioosi
Meioosi on tyyppinen solujako, joka vähentää kromosomien lukumäärää puoleen. Siten diploidisolu on jaettu neljään haploidiseen tytärsoluun. Meioosi tapahtuu itävissä soluissa ja aiheuttaa sukusoluja.
Meioosin vaiheet koostuvat kahdesta ytimen ja sytoplasman jakautumisesta, nimittäin meioosista I ja meioosi II. Meioosin i aikana kunkin homologisten kromosomien jäsenet erottuvat. Meiosis II: n aikana sisarkromatidit erilliset ja tuotetaan neljä haploidisolua.
Jokainen mitoosivaihe on jaettu profaasiin, luvattuihin, metafaasiin, anafaasiin ja tefelaasiin.
Voi palvella sinua: Bakteerisoluseinä: Ominaisuudet, biosynteesi, toiminnotMeiosis I
- Profase i. Kromosomit tiivistyvät ja kara alkaa muodostua. DNA on kaksinkertaistunut. Jokainen kromosomi koostuu sisarkromatideista yhdessä sentromeerin kanssa. Homologiset kromosomit ilmestyvät synapsen aikana, mikä mahdollistaa ristien linkin, mikä on avain erilaisten sukusolujen tuottamiseen.
- Metafaasi I. Homologisten kromosomien pari on kohdistettu metafaasilevyä pitkin. Chiasma auttaa pitämään kiinnitetyn parin. Kunkin navan Cinetocoron mikrotubulukset liittyvät homologisen kromosomin sentromeeriin.
- Anafaasi I. Cinetocoron mikrotubulukset lyhennetään ja homologiset parit erotetaan. Kopiointikunta menee solun napaan, kun taas toinen kaksoiskappale menee navan toiselle puolelle.
- TELOFASE I. Erilliset homologit muodostavat ryhmän jokaisessa solupylissä. Ydinkuori on jälleen. Citosinesis tapahtuu. Tuloksena olevilla soluilla on puolet alkuperäisten solukromosomien lukumäärästä.
Meioosi II
- Profase II. Jokaisessa solussa muodostuu uusi kara ja solukalvo katoaa.
- Metafaasi II. Karan muodostuminen on valmis. Kromosomeissa on sisarekromatideja, jotka on liitetty sentromeriin, kohdistettuna metafaasilevyä pitkin. Cinetocoron mikrotubulukset, jotka alkavat vastakkaisista pylväistä, sitoutuvat sentromeereihin.
- Anafaasi II. Mikroutkuja lyhennetään, sentromeerit on jaettu, sisarkromatidit erottuvat ja liikkuvat kohti vastakkaisia puolia.
- TELOFASE II. Ydinkääre on muodostettu neljä kromosomiryhmää: muodostuu neljä haploidisolua.
Merkitys
Joidenkin esimerkkien avulla havainnollistetaan erityyppisten solujen jakautumisen merkitys.
- Mitoosi. Solusyklillä on peruuttamattomia pisteitä (DNA: n replikaatio, sisarkromatidien erottaminen) ja kontrollipisteet (G1/S). P53 -proteiini on avain G1 -ohjauspisteeseen. Tämä proteiini havaitsee DNA -vauriot, pysäyttää solujen jakautumisen ja stimuloi vaurioita korjaavien entsyymien aktiivisuutta.
Yli 50%: lla ihmisen syöpistä p53. P53 -mutaatiot voivat aiheuttaa syöpää.
- Meioosi. Liittyy seksuaaliseen lisääntymiseen. Evolutionary -kannalta uskotaan, että seksuaalinen lisääntyminen syntyi DNA: n korjausprosessina. Siten kromosomissa tuotetut vauriot voidaan korjata homologisen kromosomin tietojen perusteella.
Uskotaan, että diploidi valtio oli väliaikainen muinaisissa organismeissa, mutta sillä alkoi olla enemmän merkitystä, kun genomista tuli suurempi. Näissä organismeissa seksuaalisella lisääntymisellä on komplementaatio, DNA: n korjaus ja geneettinen variaatio.
Viitteet
- Alberts, b., Johnson, a., Lewis, J., ja kaali. 2007. Solumolekyylin biologia. Garland Science, New York.
- Bernstein, H., Byers, G.S., Michod, R.JA. 1981. Seksuaalisen lisääntymisen kehitys: DNA: n korjauksen, komplementation ja variaation merkitys. Amerikkalainen luonnontieteilijä, 117, 537-549.
- Loodish, h., Berk, a., Zipurski, S.Lens., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Solu- ja molekyylibiologia. Pan -American Medica -toimitus, Buenos Aires.
- Raven, P.H., Johnson, G.B -., Losos, j.B -., Laulaja, S.R -. 2005 biologia. Korkeakoulu, Boston.
- Salomon, b.M., Berg, l.R -., Martin, D.W -. 2008. biologia. Thomson, USA.
- « Teeporingo -ominaisuudet, sukupuuttoon liittyvä vaara, elinympäristö
- Ricinus Commune -ominaisuudet, elinympäristö, toksisuus, käyttää »