

Glykolyysitoiminnot, entsyymit, vaiheet, tuotteet, tärkeys
- 2944
- 599
- Gustavo Runte DVM
Se Glykolyysi o Glykolyysi on tärkein glukoosikatabolismin reitti, jonka perimmäinen tavoite on saada energiaa ATP: n ja NADH: n vähentämisvoiman muodossa tästä hiilihydraatista.
Tämä reitti, jonka Gustav Embdenin ja Otto Meyerhof on täysin selvittänyt 1930 -luvulla tutkittaessa luuston lihassolujen glukoosin kulutusta, koostuu mainitun monosakkaridin täydellisestä hapettumisesta ja edustaa sinänsä anaerobista reittiä energian saamiseksi energian saamiseksi.
ATP: n molekyylirakenne, yksi glykolyyttisistä tuotteista (yhteenveto glykolyyttisestä reitistä (lähde: Tekks englanniksi Wikipedia/CC BY-SA (https: // creativecommons.Org/lisenssit/by-SA/3.0) Wikimedia Commonsin kautta) Wikimedia Commonsin kautta) Se on yksi tärkeimmistä aineenvaihduntareiteistä, koska se tapahtuu erimielisyyksillä kaikissa olemassa olevissa elävissä organismeissa, yksisoluisissa tai monisoluisissa, prokaryooteissa tai eukaryooteissa, ja ajatellaan, että se on reaktioketju, joka on hyvin säilynyt evoluutiossa luonnossa luonnossa.
Itse asiassa on joitain organismeja ja solutyyppejä, jotka riippuvat yksinomaan tästä reitistä toimimaan.
Ensinnäkin glykolyysi koostuu glukoosin, 6 hiiliatomin hapettumisesta pyruvaattiin, jolla on kolme hiiliatomia; ATP: n ja NADH: n samanaikaisen tuotannon avulla hyödyllinen soluille metabolisesta ja synteettisestä näkökulmasta.
Niissä soluissa, jotka kykenevät edelleen käsittelemään glukoosikatabolismista saatuja tuotteita, glykolyysi päättyy hiilidioksidin ja veden tuotantoon Krebs -syklin ja elektronikuljetinketjun läpi (aerobinen glykolyysi).
Kymmenen entsymaattista reaktiota tapahtuu glykolyyttisen reitin aikana, ja vaikka näiden reaktioiden säätely voi olla jotain erilaista kuin laji toiseen, säätelymekanismit ovat myös melko säilyneitä.
[TOC]
Glykolyysifunktiot
Metabolisen näkökulmasta glukoosi on yksi tärkeimmistä hiilihydraateista kaikille eläville olentoille.
Se on vakaa ja erittäin liukoinen molekyyli, joten sitä voidaan kuljettaa suhteellisen helposti koko eläimen tai kasvin kehossa, mistä se varastoidaan ja/tai saadaan sinne, missä sitä tarvitaan solupolttoaineena.
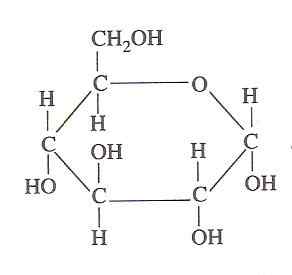 Glukoosirakenne (lähde: Oliva93/cc By-S (https: // creativecommons.Org/lisenssit/by-SA/3.0) Wikimedia Commonsin kautta)
Glukoosirakenne (lähde: Oliva93/cc By-S (https: // creativecommons.Org/lisenssit/by-SA/3.0) Wikimedia Commonsin kautta) Elävät solut hyödyntävät glukoosissa sisältävää kemiallista energiaa glykolyysin kautta, joka koostuu sarjasta erittäin kontrolloituja vaiheita, joilla tämän hiilihydraatin hapettumisesta vapautuneen energian energian hapettumisesta voidaan "vangita", joten sen merkitys sen merkitys.
Tämän reitin kautta saadaan vain energiaa (ATP) ja vähentämällä tehoa (NADH), vaan se tarjoaa sarjan aineenvaihduntavälittäjiä, jotka ovat osa muita reittejä, jotka ovat tärkeitä myös anabolisten (biosynteettisten) ja yleisen solujen toiminnan perusteella. Tässä on luettelo:
- Glukoosi 6-fosfaatti Pentotosas-fosfaattireitille (PPP, englanti Pantos Mushco -polku-A
- Maito-
- Pyruvaatti aminohapposynteesille (alaniini, lähinnä)
- Pyruvaatti trikarboksyylihapposyklille
- Fruktoosi 6-fosfaatti, glukoosi 6-fosfaatti ja dihydroksiasetonifosfaatti, jotka toimivat ”rakennuslohkoina” muilla reiteillä, kuten glykogeenisynteesi, rasvahapot, triglyseridit, nukleotidit, aminohapot jne.
Energiantuotanto
Glykolyyttisen polun tuottaman ATP: n määrä.
Aerobisten solujen suhteen glykolyysi toimii kuitenkin myös hätäenergialähteenä ja toimii "valmistelevana askeleena" ennen oksidatiivisia fosforylaatioreaktioita, jotka karakterisoivat soluja aerobisen aineenvaihdunnan kanssa.
Glykolyysiin osallistuvat entsyymit
Glykolyysi on mahdollista vain 10 entsyymin osallistumisen ansiosta, jotka katalysoivat tätä reittiä karakterisoivia reaktioita. Monet näistä entsyymeistä ovat alosteisia ja muuttavat muodon tai konformaation, kun he käyttävät katalyyttisiä toimintojaan.
On entsyymejä, jotka rikkovat ja muodostavat kovalenttisia sidoksia substraattiensa keskuudessa, ja on muitakin, jotka vaativat erityisiä kofaktoreita toimintojensa, pääasiassa metallisten ionien, käyttämiseen.
Rakenteellisesti kaikilla glykolyyttisilla entsyymeillä on keskipiste β yhdensuuntainen ympäröimä Α ja tilattu useammalla kuin yhdellä alueella. Lisäksi nämä entsyymit on karakterisoitu, koska niiden aktiiviset paikat ovat yleensä unionin alueilla verkkotunnusten välillä.
On myös tärkeää korostaa, että reitin pääasiallinen säätely kulkee entsyymien, kuten heksokinaasin, fosfrukerachinaasin, glyseraldehydi 3-fosfaattidehydrogenaasin ja pyruvaattikinaasin, kontrollin (hormonaaliset tai metaboliitit).
Voi palvella sinua: keuhkoparenchyma: kuvaus, histologia, sairaudet Glykolyyttisen polun pääasialliset säätelypisteet.Org/lisenssit/by-SA/3.0) Wikimedia Commonsin kautta)
Glykolyyttisen polun pääasialliset säätelypisteet.Org/lisenssit/by-SA/3.0) Wikimedia Commonsin kautta) 1- heksokinaasi (HK)
Glykolyysin (glukoosifosforylaatio) ensimmäistä reaktiota katalysoi heksokinaasi (HK), jonka vaikutusmekanismi näyttää koostuvan substraatin "indusoidusta säätämisestä", joka edistää ATP: n ja glukoosin ympärillä olevan entsyymin "sulkemista" (glukoosin (sulkeminen "((sulkeutuminen" (glukoosin ( hänen substraatinsa) kun he ovat liittyneet näihin.
Tarkasteltavasta organismista riippuen voi olla yksi tai useampi isoentsyymit, joiden molekyylipaino vaihtelee välillä 50 (noin 500 aminohappoa) ja 100 kDa, koska ne näyttävät olevan ryhmitelty rajojen muodossa, jonka muodostumista suosii Glukoosin, magnesiumionien ja ATP: n läsnäolo.
Heksokinaasissa on tertiäärinen rakenne, joka koostuu alfa- ja beetalevyistä, vaikka näissä entsyymeissä on monia rakenteellisia eroja.
2- Isomeraasifosfoglukoosi (PGI)
Fosforyloitu glukoosi heksakinaasin avulla isomerisoidaan 6-fosfaattifruktoosiin isomeraasifosfoglukoosin (PGI) avulla, joka tunnetaan myös nimellä glukoosi 6-fosfaatti-isomeraasi. Entsyymi ei siis poista tai lisää atomeja, vaan pikemminkin uudelleenjärjestelmä rakenteellisella tasolla.
Tämä on aktiivinen entsyymi sen dimorisessa muodossa (monomeeri painaa enemmän tai vähemmän 66 kDa) ja liittyy paitsi glykolyysiin, myös glukoneogeneesiin, kasveissa jne. Hiilihydraattien synteesissä jne.
3- fosfofrutequinaasi (PFK)
6-fosfaattifruktoosi on substraatti fosfrukerakinaasientsyymille, joka kykenee fosforylaatioon uudelleen tämä molekyyli käyttämällä ATP: tä fosforyliryhmän luovuttajana, tuottaen 1,6-biffosfaattfruktoosia.
Tämä entsyymi esiintyy bakteereissa ja nisäkkäissä homotrameerisena entsyyminä (koostuu neljästä identtisestä alayksiköstä 33 kDa bakteereille ja 85 kDa kumpikin nisäkkäissä) ja hiivoissa se on oktametri (koostuu suuremmista alayksiköistä, välillä 112 ja 118 kDa).
Se on alosterinen entsyymi, mikä tarkoittaa, että se on positiivisesti tai negatiivisesti säätelemällä mitä tahansa sen tuotteita (ADP) ja muita molekyylejä, kuten ATP ja sitraatti.
4- aldolaasi
Tunnetaan myös nimellä fruktoosi 1,6-bifosfaatti-aldolaasi, aldolaasi katalysoi fruktoosin 1,6-bifosfaatin katalyyttistä repeämää dihydroksiasetonin fosfaatissa ja glyseraldehydissä 3-fosfaatti- ja reaktioreaktio, toisin sanoen molempien sokerien yhdistäminen fruktoosin muodostumisen muodostumista varten.
Toisin sanoen tämä entsyymi leikkaa fruktoosin 1,6-bifosfaattia vain keskellä, vapauttaen kaksi 3 hiiliatomin fosforyloitua yhdistettä. Aldolaasi koostuu myös 4 identtisestä alayksiköstä, jokaisella on oma aktiivinen paikka.
Tämän entsyymin kahden luokan (I ja II) olemassaolo on määritetty, jotka eroavat katalysoivan reaktion mekanismin mukaan ja koska jotkut (ensimmäiset) esiintyvät "alemmissa" bakteereissa ja eukaryooteissa ja muissa (muut (muut (muut (muut) (muut (muut) (muut (muut) (muut (muut) (muut (muut) toinen) ovat bakteereissa, protisteissa ja metatsoissa.
”Ylivoimaisten” eukaryootien aldolaasi koostuu 40 kDa: n molekyylipainon alayksiköiden homotermaterimesta, joka muodostuu tynnyristä, joka koostuu 8 β/a -arkista.
5- triosa-fosfaatti-isomeraasi (TIM)
Kaksi triosasa fosforyloitua voidaan toistaa toistensa kanssa isomeraasitriosa-fosfaatin vaikutuksen ansiosta, mikä mahdollistaa molempien sokerien käytön koko glykolyysiin, mikä varmistaa kunkin tielle kulkevan glukoosimolekyylin täydellisen käytön.
Tätä entsyymiä on kuvattu "täydelliseksi" entsyymiksi, koska se katalysoi noin miljardia kertaa kuvatun reaktion kuin tapahtuisi ilman sen osallistumista. Sen aktiivinen paikka sijaitsee beeta-tynnyrien rakenteen keskellä, joka on ominaisuus monille glykolyyttisille entsyymeille.
Se on DiMérica -proteiini, jonka muodostuu kahdella identtisellä alayksiköllä, jotka ovat noin 27 kDa, molemmat globaalin rakenteen kanssa.
6- Glyeraldehydi 3-fosfaattidehydrogenaasi (GAPDH)
Isomeraasitriosa-fosfaatin vaikutuksella tuotettu 3-fosfaattiglyseraldehydi toimii substraattina GAPDH: lle, joka on homoterraameerinen entsyymi (34-38 kDa kutakin alayksikköä), joka sitoutuu yhteistyössä NAD+ -molekyyliin sen 4 aktiivisessa kohdassa , samoin kuin 2 fosfaatti- tai sulfaatti -ionia.
Tällä reitin kulkulla entsyymi mahdollistaa yhden sen substraatin fosforylaation, jota käytettiin epäorgaanisen fosfaatin fosforyyliryhmän luovuttajana, vähentämällä kahta NAD+ -molekyyliä ja tuotantoa 1,3-bifospphoglyssato.
7- Fosfoglyssato-kinaasi (PGK)
Fosfoglysererate-kinaasi on vastuussa yhden 1,3-bifosglyseraation fosfaattiryhmien siirtämisestä ADP. Tämä entsyymi käyttää samanlaisia mekanismia kuin heksokinaasi, koska se sulkeutuu sen substraateihin kosketukseen, suojaamalla niitä häiritseviltä vesimolekyyleiltä.
Voi palvella sinua: Aponeuroosi: Ominaisuudet, toiminnot, sijaintiTällä entsyymillä, kuten muutkin, jotka käyttävät kahta tai useampaa substraattia, on sitoutumispaikka ADP: lle ja toinen fosfaattisokerille.
Toisin kuin muut kuvatut entsyymit, tämä proteiini on 44 kDa: n monomeeri, jolla on bilobulaarinen rakenne, joka koostuu kahdesta saman kokoisesta domeenista, jotka on kytketty kapealla "uralla",.
8- Mutaasifosfoglyseraatio
3-fosfoglyseraatio muuttuu fosfaattiryhmästä hiili 2: een, molekyylin keskellä, joka edustaa strategista epävakautta, joka helpottaa ryhmän myöhempää siirtoa ATP-molekyyliin reitin viimeisessä reaktiossa.
Tätä taka -regloa katalysoi entsyymi -fosfoglysererate mutasa, dimérica -entsyymi ihmisille ja hiivoille tetrameerinen, alayksikön koko on lähellä 27 kDa.
9- enolaasi
Enolaasi katalysoi 2-fosfoglyseraation kuivumista fosfoenoliruvaatiksi, välttämätön vaihe ATP: n muodostumiseksi seuraavassa reaktiossa.
Se on dimérica -entsyymi, joka koostuu kahdesta identtisestä alayksiköstä 45 kDa. Se riippuu stabiilisuuden magnesiumioneista ja substraatin liittymiseksi tarvittavista konformaatiomuutoksista. Se on yksi monien organismien sytosolissa ilmaistuista entsyymeistä ja harjoittaa ylimääräisiä toimintoja glykolyyttisiksi.
10- Quinasa Piruvato
Toista fosforylaatiota glykolyysissä tapahtuvan substraattitasolla katalysoi kinaasi -pyruvaatti, joka vastaa fosforyyliryhmän siirrosta fosfori -fosfoenoliruvaatti ADP: hen ja pyruvaatin tuotantoon.
Tämä entsyymi on monimutkaisempi kuin mikään muu glykolyyttistä entsyymejä ja nisäkkäissä se on homotramalentsyymi (57 kDa/alayksikkö). Selkärankaisilla on vähintään 4 isoentsyymiä: L (maksassa), R (erytrosyyteissä), M1 (lihaksissa ja aivoissa) ja M2 (sikiön kudos ja aikuiset kudokset).
Glykolyysivaiheet (askel askeleelta)
Glykolyyttinen reitti koostuu kymmenestä peräkkäisestä vaiheesta ja alkaa glukoosimolekyylistä. Prosessin aikana glukoosimolekyyli "aktivoidaan" tai "valmistettu" lisäämällä kaksi fosfaattia, sijoittamalla kaksi ATP -molekyyliä.
Myöhemmin se "leikataan" kahdessa fragmentissa ja sitä muutetaan lopulta kemiallisesti pari kertaa syntetisoimalla tiellä neljä ATP -molekyyliä, joten reitin nettovahvistus vastaa kahta ATP -molekyyliä.
Edellä mainitusta voidaan päätellä, että reitti on jaettu energia "sijoitus" -vaiheeseen, perustavanlaatuiseen glukoosimolekyylin täydelliseen hapettumiseen ja toiseen energian ”voiton” vaiheeseen, jossa alun perin käytetty energia täydennetään ja kaksi ATP -verkkoa molekyylit.
- Energiainvestointivaihe
1- Glykolyyttisen reitin ensimmäinen vaihe koostuu heksakinaasin (HK) välittämän glukoosin fosforylaatiosta, jolle entsyymi käyttää ATP-molekyyliä jokaiselle fosforyloidulle glukoosimolekyylille, joka on fosforyloitu. Se on peruuttamaton reaktio ja riippuu magnesiumionien läsnäolosta (mg2+):
Glukoosi + ATP → Glukoosi 6-fosfaatti + ADP
Näin tuotettu 2- 6-fosfaattiglukoosi isomerisoidaan 6-fosfaattifruktoosiin isomeraasifosfoglukoosientsyymin (PGI) (PGI) vaikutuksen ansiosta (PGI) (PGI). Tämä on palautuva reaktio, eikä se tarkoita ylimääräisiä energiamenoja:
Glukoosi 6-fosfaatti → Fruktoosi 6-fosfaatti
3- Myöhemmin toinen energiainvestointivaihe merkitsee fruktoosin 6-fosfaatin fosforylaatiota 1,6-bifosfaattfruktoosin muodostamiseksi. Tätä reaktiota katalysoi entsyymi fosfofrutequinaasi-1 (PFK-1). Kuten reitin ensimmäinen vaihe, fosfaattiryhmän luovuttaja molekyyli on ATP ja se on myös peruuttamaton reaktio.
Fruktoosi 6-fosfaatti + ATP → Fruktoosi 1,6-bifosfaatti + ADP
4- Tässä glykolyysin vaiheessa fruktoosin 1,6-bifosfaatin katalyyttinen repeämä dihydroksiasetonfosfaatissa (DHAP) -keosissa ja glyseraldehydi 3-fosfaatissa (aukko) Aldosa A-aldosa. Tätä aldoolista kondensaatiota katalysoi entsyymi -aldolaasi ja se on palautuva prosessi.
Fruktoosi 1,6-biffosfaatti → dihydroksiasetonifosfaatti + glyseraldehydi 3-fosfaatti
5- Energiainvestointivaiheen viimeinen reaktio koostuu triosas-DHAP-fosfaatin ja aukkojen katalysoimasta triosa-fosfaatti-isomeraasi (TIM) -entsyymistä, tosiasia, joka ei vaadi ylimääräistä energiaa ja joka on myös palautuva prosessi.
Dihydroksiasetonifosfaatti ↔ glyseraldehydi 3-fosfaatti
- Energian voittovaihe
6-3-fosfaattiglyseraldehydiä käytetään "alavirtaan" glykolyyttisellä reitillä substraattina hapettumisreaktiolle ja toiselle fosforylaatiolle, jota katalysoi sama entsyymi, glykaraldehydi 3-fosfaattihydrogenaasi (GAPDH).
Entsyymi katalysoi molekyylin hiili-C1: n hapettumista karboksyylihappoa ja tämän fosforylaatio samassa paikassa, mikä tuottaa 1,3-bifoscicerea. Reaktion aikana käytetään 2 NAD+ -molekyyliä glukoosimolekyyliä kohti ja käytetään 2 epäorgaanista fosfaattimolekyyliä.
Voi palvella sinua: henkitorvi2Glyeraldehydi 3-fosfaatti + 2Nad + + 2Pi → 2 (1,3-bifosfoglyssato) + 2NADH + 2H
Aerobisissa organismeissa jokainen tällä tavalla tuotettu NADH kulkee elektronikuljetinketjun läpi toimimaan substraattina 6 ATP.
7- Tämä on ATP: n synteesin ensimmäinen vaihe glykolyysissä ja merkitsee kinaasifosfoglyssato (PGK): n vaikutusta 1,3-bifosfoglyseraatissa, siirtämällä fosforyyliryhmää (fosforylaatio substraattitasolla) tästä molekyylistä molekyylille, saaden 2ATPP: n 2ATPP: n ja 2 molekyyliä 3-fosfoglyseraattia (3pg) jokaiselle glukoosimolekyylille.
2 (1,3-bifosfoglyssato) + 2ADP → 2 (3-fosfoglyseraatti) + 2ATP
8-3-fosfoglyseratio toimii substraattina fosfoglyssato-mutasaa (PGM), mikä tekee siitä 2-fosfoglyseraation siirtämällä hiili 3-hiili 2: n fosforyyliryhmää, joka on palautuva ja riippuvainen siitä Magnesiumioonit (mg+2).
2 (3-fosfoglyserera) → 2 (2-fosfoglyseraatio)
9- Entsyymi 2-fosfoglyseraattidehydraatissa ja tuottaa fosfoenoliruvaattia (PEP) reaktion avulla, joka ei ansaitse lisäenergian lisäämistä ja jonka tavoitteena on tuottaa korkean energian yhdiste, joka kykenee luovuttamaan fosforyyliryhmänsä seuraavassa reaktiossa.
2 (2-fosfoglyserera) → 2Fosfoenolpiruvate
10- Fosfoenoliruvaatti on pyruvaattikinaasi (PYK) -entsyymin substraatti, joka on vastuussa fosforyyliryhmän siirrosta tässä molekyylissä kohti ADP.
Reaktiossa 2ATP- ja 2 pyruvaattimolekyyliä tuotetaan jokaiselle glukoosille ja kaliumin ja magnesiumin läsnäolo ionisessa muodossa on välttämätöntä.
2Fospo Elplivato + 2ADP → 2Pyruvate + 2ATP
Glykolyysin netto suorituskyky tällä tavalla koostuu 2ATP: stä ja 2Nad+: sta jokaisesta reitille kulkevasta glukoosimolekyylistä.
Jos nämä ovat soluja, joilla on aerobinen aineenvaihdunta, glukoosimolekyylin kokonais hajoaminen tuottaa 30–32 ATP: tä Krebs -syklin ja elektronien kuljetusketjun kautta.
Glykolyysituotteet
Glykolyysin yleinen reaktio on seuraava:
Glukoosi + 2Nad + + 2ADP + 2Pi → 2Pyruvate + 2ATP + 2NADH + 2H+
Siksi, jos sitä analysoidaan lyhyesti, se voisi varmistaa, että glykolyyttisen reitin päätuotteet ovat pyruvaatti, ATP, NADH ja H.
Kunkin reaktiovälittäjän metabolinen kohde riippuu kuitenkin suurelta osin solujen tarpeista, minkä vuoksi kaikkia välittäjiä voidaan pitää reaktiotuotteina, jotta ne voivat lainaamaan niitä seuraavasti:
- 6-fosfaatti
- 6-fosfaatti
- Fruktoosi 1,6-bifosfaatti
- Dihydroksiasetonifosfaatti ja glyseraldehydi 3-fosfaatti
- 1,3-bifosfoglyssato
- 3-fosfoglyseraatio ja 2-fosfoglyseraatio
- Fosfoenoliruvaatti ja pyruvaatti
Merkitys
Vaikka glykolyysi sinänsä (voisi puhua anaerobisesta glykolyysistä) tuottaa vain noin 5% ATP: stä, joka voidaan erottaa glukoosin aerobisesta katabolismista, tämä metabolinen reitti on välttämätöntä monista syistä:
- Se toimii "nopeana" energialähteenä, etenkin tilanteissa, joissa eläimen on jätettävä lepotila nopeasti, jolle aerobiset hapettumisprosessit eivät olisi tarpeeksi nopeita.
- Esimerkiksi ihmiskehon luuston "valkoiset" lihaskuidut ovat nopeita supistumiskuituja ja riippuvat anaerobisesta glykolyysistä toimimaan.
- Kun solu tarvitsee jostain syystä glykolyyttistä.
- Monet solut ovat riippuvaisia glukoosista energian lähteenä glykolyyttisellä reitillä, mukaan lukien punasolut, Sisäisten organelien ja silmälasien (etenkin sarveiskalvon) puuttuvat, joilla ei ole suurta mitokondriatiheyttä.
Viitteet
- Canback, b., Andersson, S. G. JA., & Kurland, c. G. (2002). Glykolyyttisten entsyymien globaali fylogenia. Kansallisen tiedeakatemian julkaisut, 99 (9), 6097-6102.
- Chaudhry R, Varacallo M. Biokemia, glykolyysi. [Päivitetty 2019 AP 21]. Julkaisussa: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2020 tammikuu-. Saatavana osoitteesta: https: // www.NCBI.Nlm.NIH.Gov/Books/NBK482303/
- Fothergill-gilmore, l. -Lla., & Michels, P. -Lla. (1993). Glykolyysin kehitys. Biofysiikan ja molekyylibiologian eteneminen, 59 (2), 105-235.
- Kim, J. W -., & Dang, c. V. (2005). Glykolyyttisten entsyymien monipuoliset roolit. Biokemiallisten tieteiden trendit, 30 (3), 142-150.
- Kumari, a. (2017). Makea biokemia: Mnemoniikan rakenteiden, syklien ja polkujen muistaminen. Akateeminen lehdistö.
- Li, x. B -., Gu, J. D -d., & Zhou, Q. H. (2015). Katsaus aerobiseen glykolyysiin ja sen keskeisiin entsyymit-uusiin kohteisiin keuhkosyöpähoidossa. Rintakehän syöpä, 6 (1), 17-24.
- « Kasvissyöjäeläinten ominaisuudet, tyypit, esimerkit
- Tupakan mosaiikkiviruksen ominaisuudet, rakenne, replikaatio »