

Microcuerpos -ominaisuudet, toiminnot ja esimerkit
- 1244
- 132
- Dr. Travis Reichert
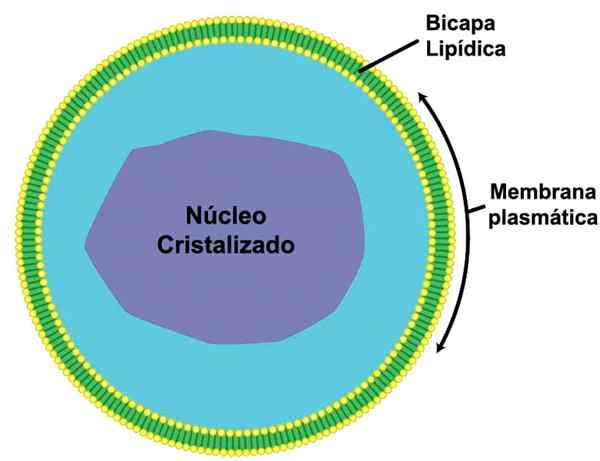
Se Mikrovirrat Ne muodostavat luokan sytoplasmisia organeleja, joita ympäröi yksinkertainen kalvo ja jotka sisältävät hienon matriisin, jolla on muuttuva ulkonäkö amorfisen, fibrillaarisen tai rakeisen muodon välillä. Mikrokanassa esiintyy joskus erilainen keskuksen tai ytimen, jolla on suurempi elektroninen tiheys ja kiteinen asennus.
Näissä organelissa on useita entsyymejä, joista osa on hapettumisfunktio (kuten katalaasi), jotka osallistuvat joidenkin ravinteiden hapettumiseen. Esimerkiksi peroksisomit hajottavat vetyperoksidin (H2JOMPIKUMPI2-A.
Peroksisomin graafinen esitys.Lähde: Rock 'N Roll [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]]
Niitä löytyy eukaryoottisista soluista ja ne ovat peräisin sisällyttämällä proteiineja ja lipidejä sytoplasmasta ja ympäröivistä kalvoyksiköistä.
[TOC]
Ominaisuudet
Microcuerpos voidaan määritellä vesikkeleiksi yksinkertaisella kalvolla. Näiden organelien halkaisija on välillä 0,1 - 1,5 um. Niillä on munanmuoto ja joissain tapauksissa pyöreä, rakeinen ulkonäkö. Joskus marginaalinen plakki voidaan esitellä organellin keskellä, mikä antaa sille tietyn muodon.
Nämä pienet rakenteet löydettiin äskettäin ja karakterisoitiin morfologisesti ja biokemiallisesti elektronisen mikroskopian kehityksen ansiosta.
Eläinsoluissa ne sijaitsevat lähellä mitokondrioita, ovat aina paljon pienempiä kuin nämä. Mikrokana liittyy myös alueellisesti sileään endoplasmiseen retikulumiin.
Mikrovirtauskalvo koostuu Porinasta ja on ohuempi kuin muiden organelien, kuten lysosomien, ja joissain tapauksissa pienten molekyylien läpäisevää (kuten maksasoluissa peroksisomeissa).
Mikrokanan matriisi on yleensä rakeinen ja joissain tapauksissa homogeeninen, elektroninen tiheys on yleensä tasainen ja haarautuneilla filamenteilla tai lyhyillä fibrilleillä. Entsyymien sisältämisen lisäksi löydämme paljon fosfolipidejä.
Funktiot
Eläinsoluissa
Microcuerpos osallistuu moniin biokemiallisiin reaktioihin. Ne voivat liikkua solussa paikkaan, jossa niiden toiminnot vaaditaan. Eläinsoluissa ne liikkuvat mikrotubulusten ja kasvisolujen välillä, ne liikkuvat mikrofilamentteja pitkin.
Voi palvella sinua: erytroblastit: mitkä ovat erytropoieSis, liittyvät patologiatNe toimivat saamana vesikkelia erilaisista aineenvaihduntareitteistä, jotka toimivat niiden kuljettamisessa, ja heissä tapahtuu jonkin verran aineenvaihduntaa.
Peroksisomit tuottavat h2JOMPIKUMPI2 vähentämisestä tai2 Alkoholilla ja pitkän ketjun rasvahapoilla. Tämä peroksidi on erittäin reaktiivinen aine ja sitä käytetään muiden aineiden entsymaattisessa hapettumisessa. Peroksisomit täyttävät H: n hapettumisen solukomponenttien suojaamisen tärkeän toiminnan H: n avulla2JOMPIKUMPI2 halventamalla sitä sisälle.
P-hapettumisessa peroksisoomat ovat hyvin lähellä lipidejä ja mitokondrioita. Ne sisältävät rasvan hapettumiseen osallistuvia entsyymejä, kuten katalaasi, liosa ja paha isoklaattiset syntaasi. Ne sisältävät myös lipaaseja, jotka hajottavat rasvat, jotka on varastoitu rasvahappoketjuihinsa.
Peroksisoomat syntetisoivat myös sappisuoloja, jotka auttavat lipidimateriaalin sulamista ja imeytymistä.
Kasvisoluissa
Kasveista löydämme peroksisomit ja glioksisomit. Nämä mikrokuvat ovat rakenteellisesti samat, vaikka niillä on erilaisia fysiologisia toimintoja. Peroksisomeja löytyy verisuonikasvien lehdistä ja niihin liittyy kloroplastien. Niissä glykolyyttihapon hapettuminen tapahtuu CO: n kiinnittämisen aikana2.
Glioksisomaa löytyy runsaasti siementen itävyyden aikana, jotka ylläpitävät lipidireservyä. Glioksylaattisykliin osallistuvat entsyymit, joissa lipidien muuntaminen hiilihydraateiksi tapahtuu, niitä löytyy näistä mikrokoruista.
Fotosynteettisten koneiden paljastumisen jälkeen hiilihydraatit muodostuvat valokuvien respiraatioreitin kautta peroksisomeissa, missä kadonnut hiili otetaan oman O: n liiton jälkeen2 Rubiscolle.
Mikrokerrat sisältävät kataloja ja muita flaviiniriippuvaisia oksidaaseja. Flaviiniin kytkettyjen oksidaasien substraattien hapettumiseen liittyy hapenkeräys ja siitä johtuva H: n muodostuminen2JOMPIKUMPI2. Tätä peroksidia hajoaa katalaasin vaikutuksesta, tuottaen vettä ja happea.
Nämä organelit edistävät happea keräilyä solun avulla. Vaikka toisin kuin mitokondriat, ne eivät sisällä elektronisia kuljetusketjuja tai muuta energiaa vaativaa järjestelmää (ATP).
Se voi palvella sinua: megakariosyyttit: ominaisuudet, rakenne, muodostuminen, kypsyminenEsimerkit
Vaikka mikrokorut ovat hyvin samankaltaisia toistensa kanssa niiden rakenteen suhteen, ne on erotettu erityyppisiksi, suoritettujen fysiologisten ja metabolisten toimintojen mukaan.
Peroksisomit
Peroksisoomat ovat mikrokaanaa, jota ympäröivät halkaisijaltaan noin 0,5 um: n kalvo, jolla on erilaisia hapettumisentsyymejä, kuten katalaasi, D-aminohappo-oksidaasi, Urato-oksidaasi. Nämä organelit muodostuvat endoplasmisista retikulum -projektioista.
Peroksisoomia löytyy suuresta määrästä selkärankaisten soluja ja kudoksia. Nisäkkäissä löytyy maksa- ja munuaisten soluista. Aikuisissa rotilla maksasoluissa on havaittu, että mikrokourut ovat välillä 1 - 2% sytoplasmisen kokonaismäärästä.
Löydät mikrokuoret useista nisäkkäiden kudoksista, vaikka ne eroavat maksasta ja munuaisista löydetyistä peroksisomeista katalaasiproteiinin esittämiseksi vähemmän määrään ja puuttuvat suurin osa näissä maksasolun organeleissa olevista oksidaaseista.
Joissakin protisteissa on myös tärkeitä määriä, kuten Tetrahymya pyriformis.
Maksasoluissa, munuaisissa ja muissa prostistikudoksissa ja organismeissa esiintyneet peroksisomit eroavat toisistaan koostumuksessa ja joidenkin toimintojensa aikana.
Maksa
Maksasoluissa mikrokerrat koostuvat enimmäkseen katalaasista, joka muodostaa noin 40% näiden organelien kokonaisproteiineista. Muita oksidaaseja, kuten kumproteiineja, oksidaasi-Uatoa, flavoproteiineja ja D-aminohappo-oksidaasia, löytyy maksaperoksisomeista.
Näiden peroksisomien kalvoa jatketaan yleensä sileän endoplasmisen retikulumin kanssa liitteen tyyppisen projektion kautta. Matriisilla on kohtalainen elektronitiheys ja rakenne amorfisen ja rakeisen välillä. Sen keskuksessa on suuri elektroninen tiheys ja sillä on poliputkinen rakenne.
Munuaiset
Hiirillä ja rotilla munuaisten soluista löydetyllä mikrokanalla on rakenteelliset ja biokemialliset ominaisuudet hyvin samankaltaiset kuin elävät soluperoksisomit.
Voi palvella sinua: Langerhans -solut: Ominaisuudet, morfologia, toiminnotNäiden organelien proteiini- ja lipidikomponentit vastaavat maksasolujen kanssa. Rotan munuaisten peroksisomeissa oksidaasi -urato ei kuitenkaan ole ja katalaasi ei ole suurina määrinä. Munuaishiirisoluissa peroksisomista puuttuu elektroninen tiheyskeskus.
Tetrahymya pyriformis
Peroksisoomien esiintyminen erilaisissa protisteissa, kuten T. pyriformis, Katalasentsyymien, D-aminohappo-oksidaasin ja L-a-hydroksihappo-oksidaasin aktiivisuuden havaitsemiseksi.
Glioksisomit
Joissakin kasveissa on erikoistuneita peroksisomia, joissa glioksylaattireitin reaktiot tapahtuvat. Näitä organeleja kutsuttiin glioksisomeiksi, koska ne kantavat entsyymejä ja myös tämän aineenvaihdunnan reitin reaktiot.
Glykosomit
Ne ovat pieniä organeleja, jotka suorittavat glykolyysiä joissain alkueläimissä, kuten Trypanosoma SPP. Glykolyysin alkuvaiheisiin osallistuvat entsyymit liittyvät tähän organelli (HK, fosfoglukoosi -isomeraasi, PFK, ALD, TIM, Kinaasiglyseroli, GAPDH ja PGK).
Nämä ovat homogeenisia ja halkaisija on noin 0,3 um. Noin 18 tähän mikropuheluun liittyvää entsyymiä on löydetty.
Viitteet
- Cruz-Reyes, a., & Camargo-Camargo, B. (2000). Parasitologian ja siihen liittyvien tieteiden termien sanasto. Ja Valdes.
- Duve, c. -Lla. B -. P., & Baudhuin, P. (1966). Peroksisomit (mikrokehykset ja niihin liittyvät kumppanit). Fysiologiset arvostelut, 46(2), 323-357.
- Hruban, Z., & RechCligl, M. (2013). Mikrokehykset ja niihin liittyvät hiukkaset: morfologia, biokemia ja fysiologia (Vol. 1). Akateeminen lehdistö.
- Madigan, m. T., Markinko, J. M. & Parker, J. (2004). Brock: Mikro -organismbiologia. Pearson -koulutus.
- Nelson, D. Lens., & Cox, M. M. (2006). Lehninger -biokemian periaatteet 4. painos. Ed Omega. Barcelona.
- Smith, H., & Smith, H. (Toim.-A. (1977). Kasvisolujen molekyylibiologia (Vol. 14). Kalifornian yliopisto.
- Voet, D., & Voet, J. G. (2006). Biokemia. Ed. Pan -American Medical.
- Wayne, r. JOMPIKUMPI. (2009). Kasvien solubiologia: Tähtitieteestä eläintieteisiin. Akateeminen lehdistö.
- « Endosytoosityypit ja niiden ominaisuudet, toiminnot, esimerkit
- Ovoviviparos -ominaisuudet ja esimerkit »