

Mitoosifaasit, ominaisuudet, toiminnot ja organismit
- 3445
- 307
- Alonzo Kirlin
Se mitoosi Se on solunjakoprosessi, jossa solu tuottaa geneettisesti identtisiä tytärsoluja; Jokaiselle solulle kaksi "tytärtä" syntyy samalla kromosomaalisella kuormalla. Tämä jako suoritetaan eukaryoottisten organismien somaattisissa soluissa.
Tämä prosessi on yksi eukaryoottisten organismien solusyklin vaiheista, jotka ymmärretään 4 vaiheessa: S (DNA -synteesi), M (solunjako), G1 ja G2 (välivaiheet, joissa mRNA: t ja proteiinit esiintyvät). Yhdessä vaiheet G1, G2 ja S pidetään käyttöliittymänä. Ydin- ja sytoplasminen jako (mitoosi ja sytokiinit) muodostavat solusyklin viimeisen vaiheen.
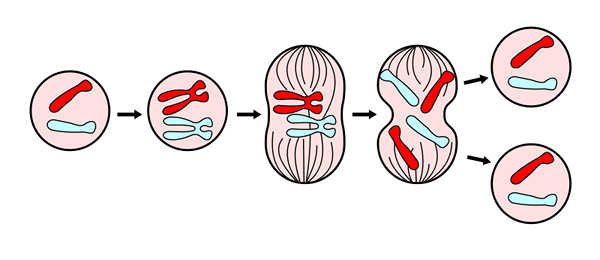
Mydoosin yleiskatsaus. Lähde: Viewaprabha [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Molekyylitasolla myitoosi aloitetaan kinaasin (proteiinin) aktivaatiolla, jota kutsutaan MPF: ksi (kypsymistä edistävä tekijä) ja siitä johtuva merkittävän määrän solukomponenttien komponenttien fosforylaatio. Jälkimmäinen antaa solulle mahdollisuuden esittää jakoprosessin toteuttamiseksi tarvittavat morfologiset muutokset.
Mitoosi on aseksuaalinen prosessi, koska progenitorisolulla ja sen tyttärillä on täsmälleen sama geneettinen tieto. Nämä solut tunnetaan diploidina kantamalla koko kromosomaalinen kuorma (2N).
Meioosi puolestaan on solujen jakautumisprosessi, joka johtaa seksuaaliseen lisääntymiseen. Tässä prosessissa diploidinen kantasolu toistaa kromosomit ja jakaa sitten kahdesti peräkkäin (toistamatta sen geneettistä tietoa). Lopuksi, 4 tytärsolua syntyy vain puolet kromosomaalisesta kuormasta, joita kutsutaan haploideiksi (n).
[TOC]
Mitoosin yleisyys
Mitoosi yksisoluisissa organismeissa tuottaa yleensä tytärsoluja, jotka ovat hyvin samanlaisia kuin sen vanhempansa. Sitä vastoin monisoluisten olentojen kehittymisen aikana tämä prosessi voi aiheuttaa kahta solua, joilla on joitain erilaisia ominaisuuksia (huolimatta geneettisesti identtisistä).
Tämä solujen erilaistuminen aiheuttaa erilaisia solutyyppejä, jotka muodostavat monisoluiset organismit.
Organismin elämän aikana solusykli tapahtuu jatkuvasti muodostaen jatkuvasti uusia soluja, jotka puolestaan kasvavat ja valmistautuvat jakamiseen myitoosin kautta.
Solujen kasvua ja jakautumista säätelevät mekanismit, kuten apoptoosi (ohjelmoitu solukuolema), jotka sallivat tasapainon ylläpitämisen, välttäen kudosten ylimääräistä kasvua. Tällä tavoin varmistetaan, että vialliset solut korvataan uusilla soluilla, organismin vaatimusten ja tarpeiden mukaan.
Mikä merkitys tällä prosessilla on?
Kyky lisääntyä on yksi kaikkien organismien tärkeimmistä ominaisuuksista (yksisoluisesta monisoluiseen) ja soluihin, jotka säveltävät sen. Tämä laatu mahdollistaa geneettisen tietosi jatkuvuuden varmistamisen.
Mitoosin ja meioosin prosessien ymmärtämisellä on ollut perustavanlaatuinen rooli organismien kiehtovien soluominaisuuksien ymmärtämisessä. Esimerkiksi kromosomien lukumäärän pitäminen solusta toiseen yksilön sisällä ja saman lajin yksilöiden välillä.
Kun kärsimme jonkin tyyppisiä leikkauksia tai haavoja ihollamme, havaitsemme, kuinka muutaman päivän kuluessa vaurioitunut iho otetaan talteen. Tämä tapahtuu myitoosiprosessin ansiosta.
Vaiheet ja sen ominaisuudet
Yleensä mytoosi seuraa samaa prosessisekvenssiä (vaiheita) kaikissa eukaryoottisoluissa. Näissä vaiheissa solussa tapahtuu monia morfologisia muutoksia. Niistä kromosomien kondensoituminen, ydinkalvon repeämä, solun erottaminen solunulkoisesta matriisista ja muista soluista sekä sytoplasman jakautuminen.
Joissakin tapauksissa ydinjakautumista ja sytoplasmista jakautumista pidetään eri vaiheina (mitoosi ja sytokiinit, vastaavasti).
Prosessin paremman tutkimuksen ja ymmärryksen saavuttamiseksi on nimetty kuusi (6) vaihetta, joita kutsutaan: propase, luvattu, metafaasi, anafaasi ja tephaasi, ottaen sitten sytokiinit kuudentena vaiheena, joka alkaa kehittyä anafaasin aikana.
 Telophaasi on mitoosin viimeinen vaihe. Otettu https: // commons.Wikimedia.Org/wiki/tiedosto: mitosepanel.JPG. Via Wikimedia Commons
Telophaasi on mitoosin viimeinen vaihe. Otettu https: // commons.Wikimedia.Org/wiki/tiedosto: mitosepanel.JPG. Via Wikimedia Commons Näitä vaiheita on tutkittu 1800 -luvulta lähtien valomikroskoopin kautta, joten nykyään ne ovat helposti tunnistettavissa solun morfologisten ominaisuuksien, kuten kromosomaalisen kondensaation ja mitoottisen karan muodostumisen mukaan.
Profaatti
 Profaatti. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons
Profaatti. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons Profase on ensimmäinen näkyvä solujen jakautumismuoto. Tässä vaiheessa voit nähdä kromosomien esiintymisen erotettavina muodoina kromatiinin asteittaisesta tiivistyksestä johtuen. Tämä kromosomien kondensoituminen alkaa Histona H1 -molekyylien fosforylaatiolla MPF -kinaasin avulla.
Kondensaatioprosessi koostuu supistumisesta ja siksi kromosomien suuruuden vähentämisestä. Tämä johtuu kromatiinikuitujen liikkumisesta, mikä tuottaa helpommin siirtymättömiä rakenteita (mitoottisia kromosomeja).
Aikaisemmin solusyklin ajanjakson aikana kopioidut kromosomit hankkivat kaksinkertaisen filamentin ulkonäön, nimeltään sisarkromatidit, nämä filamentit pysyvät yhtenäisinä Centromero -nimisen alueen kautta. Tässä vaiheessa myös nukleolit katoavat.
Se voi palvella sinua: Kromafiinisolut: Ominaisuudet, histologia, toiminnotMitoottinen karan muodostuminen
Kirjoittanut Silvia3 [GFDL (http: // www.GNU.Org/copyleft/fdl.html) tai cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons Profaasin aikana mitoottinen kara, joka koostuu mikrotubuluksista ja proteiineista, jotka muodostavat kuitujoukon.
Kun kara muodostuu, sytoskeleton mikrotubulukset (deaktivoimalla niiden rakennetta ylläpitäviä proteiineja) ovat hämärtyneet, mikä tarjoaa tarvittavan materiaalin mainitun mitoottisen karan muodostumiseen.
Keskustat (organeli ilman kalvoa, toiminnallinen solusyklissä), kaksoiskappale rajapinnalla, toimii karan mikrotubulusten kokoonpanoyksikkönä. Eläinsoluissa keskitetyllä on keskustassa, pari keskipisteitä; Mutta näitä ei ole useimmissa kasvisoluissa.
Kaksoiskappaleet, ne alkavat erottaa toisistaan, kun taas karan mikrotubulukset on koottu jokaisessa heistä, alkaen siirtyä solun vastakkaisiin päihin.
Profaasin lopussa ydinkokouksen repeämä alkaa erillisissä prosesseissa: Ydinkuokosten MAS -valtakunta, ydinlevy ja ydinmembraanit. Tämän tauon avulla mitoottiset karat ja kromosomit voivat alkaa olla vuorovaikutuksessa.
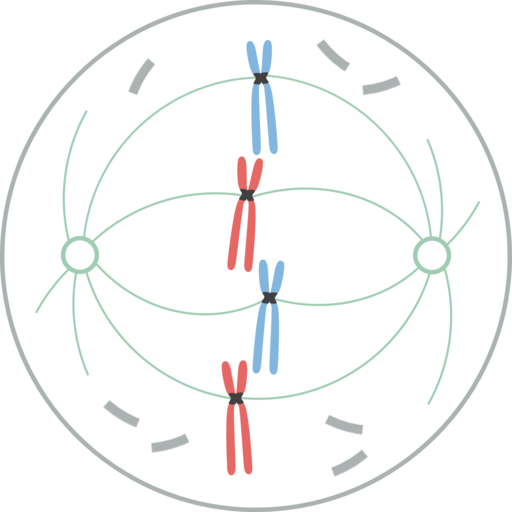
Lupaus
Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Tässä vaiheessa ydinkuori on ollut täysin pirstoutunut, joten karan mikrotubulukset tunkeutuvat tähän alueeseen vuorovaikutuksessa kromosomien kanssa. Nämä kaksi keskutusta ovat eronneet, kukin mitoottisen karan napoissa solujen vastakkaisissa ääripäissä.
Nyt mitoottinen kara sisältää mikrotubulukset (jotka ulottuvat jokaisesta keskityksestä solun keskustaan), keskukset ja parin Osteres (rakenteet, joissa on lyhytaikaisten mikrotubulusten säteittäinen jakautuminen, jotka on otettu käyttöön jokaisesta keskityksestä).
Kromatidit kehittivät jokaisen, erikoistuneen proteiinirakenteen, nimeltään Cinetocoro, joka sijaitsee sentromeerissä. Nämä kneecorot sijaitsevat vastakkaisiin suuntiin ja joitain mikrotubuluksia on kiinnitetty, nimeltään CinnetOCoron mikrotubulukset.
Nämä Cinetocoroon kiinnitetyt mikrotubulukset alkavat siirtää kromosomia lopusta, jonka lopusta ne ulottuvat; Jotkut navasta ja toiset vastakkaisesta navasta. Tämä luo "vedä ja kutistuvan" -vaikutuksen, joka stabiloinnissa antaa kromosomin päättyä solun päiden välillä.
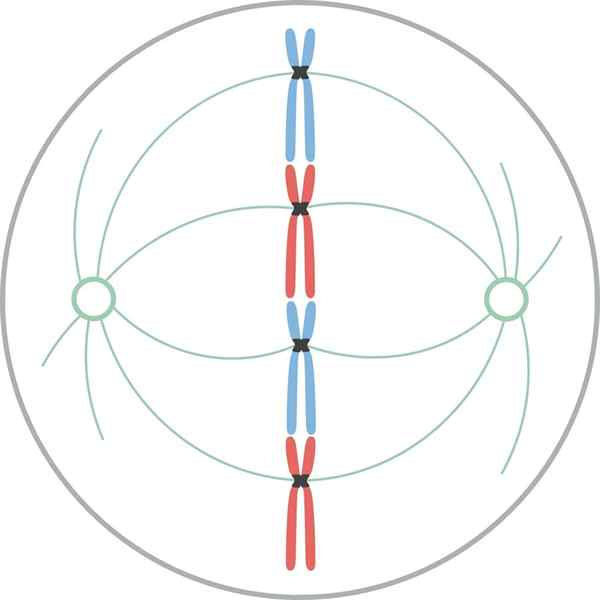
Metafaasi
Kromosomit kohdistuvat solun päiväntasaajan levyyn myyttisen metafaasin aikana Metafaasissa keskukset sijaitsevat solujen vastakkaisissa päissä. Karalla on selkeä rakenne, jonka keskellä kromosomit sijaitsevat. Näiden kromosomien sentromeerit on kiinnitetty kuiduihin ja kohdistetaan kuvitteelliseen tasoon, jota kutsutaan metafaasiseksi plakkille.
Kromatidi cipNetOCoros on edelleen kiinnitetty Cinetocoron mikrotubuluksiin. Mikrotubulukset, jotka eivät kiinnitä kneecorosia ja ulottuvat karan vastakkaisista napoista, ovat nyt vuorovaikutuksessa keskenään. Tässä vaiheessa Osteresin mikrotubulukset ovat kosketuksissa plasmamembraanin kanssa.
Tämä mikrotubulusten kasvu ja vuorovaikutus täydentää mitoottisen karan rakennetta ja antaa ulkonäön "lintuhäkki".
Morfologisesti tämä vaihe on se, joka näyttää vähemmän muutoksista, joten sitä pidettiin lepovaiheena. Vaikka ne eivät ole helposti tuntuvia, siinä tapahtuu monia tärkeitä prosesseja, sen lisäksi, että mitoosi on pisin.
Anafasi
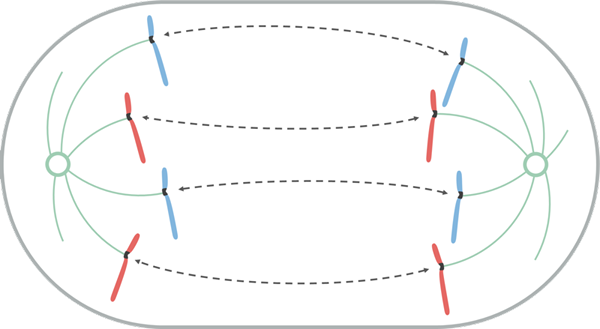 Lähde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons
Lähde: Leomonaci98 [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons Anafaasin aikana jokainen kromatidipari alkaa erottua (inaktivoimalla proteiineja, jotka pitävät niitä yhdessä). Erilliset kromosomit liikkuvat kohti solun vastakkaisia päätä.
Tämä muuttoliike johtuu siitä, että Catocoro de acortanin mikrotubulukset tuottavat "veto" -tehosteen, joka saa jokaisen kromosomin liikkeen sen centromerosta. Kromosomissa olevan sentromeerin sijainnista riippuen se voi viedä sen siirtymän aikana tietty muoto, kuten V tai J.
Mikrotubuluksia, joita ei kiinnitetä kneecoroon, kasvavat ja pidentävät tubuliiniarvikkeiden (proteiinia) ja niiden liikkuvien motoristen proteiinien vaikutusta, jolloin niiden välinen kosketus pysäyttää. Kun he siirtyvät pois toisistaan, myös karapylväät tekevät sen, pidentäen solua.
Tämän vaiheen lopussa kromosomiryhmät sijaitsevat mitoottisen karan vastakkaisissa päissä, joten solun jokainen pää on täydellisellä ja vastaavalla kromosomisarjalla.
Telfaasi
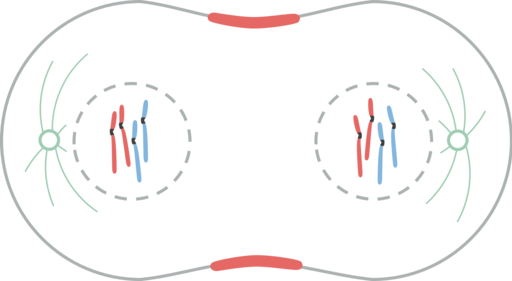 Telfaasi. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)]
Telfaasi. Leomonaci98 [cc by-sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Telophaase on ydinjaon viimeinen vaihe. Cinetocoro -mikrotubulukset hajoavat, kun taas polaariset mikrotubulukset pidennetään edelleen.
Ydinmembraani alkaa muodostua kunkin kromosomin pelin ympärille käyttämällä emosolun ydinkääreitä, jotka olivat kuin sytoplasman rakkuloita.
Tässä vaiheessa solujen pylväissä olevia kromosomeja lannistuu kokonaan histonimolekyylien defosforylaation vuoksi (H1). Ylimembraanin elementtien muodostuminen ohjaavat useat mekanismit.
Anafaasin aikana monet fosforyloituneista proteiineista alkoivat defosforoida profaasissa. Tämä mahdollistaa telfaasin alkamisen, ydinvesikkeleet alkavat koota uudelleen, liittyy kromosomien pintaan.
Voi palvella sinua: chondrosyytit: ominaisuudet, histologia, toiminnot, viljelyToisaalta ydinhuokos on uudelleenkokoonpano, joka mahdollistaa ydinproteiinien pumppaamisen. Ydinlaminaproteiinit defosforyloidaan, jotta ne voidaan yhdistää uudelleen, mainitun ydinlaaminan muodostumisen loppuun saattamiseksi.
Lopuksi, kun kromosomit ovat täysin lannistuneet, RNA: n synteesi käynnistetään uudelleen, muodostaen ytimen uudelleen ja siten täydentämällä tytärsolujen uusien rajapinnan ytimien muodostumista.
Sytokiinia

Sytocinesit otetaan erillään ydinosastosta ja yleensä tyypillisissä soluissa sytoplasmisen jakoprosessi liittyy jokaiseen myitoosiin, joka alkaa anafaasista. Useat tutkimukset ovat osoittaneet, että joissakin alkioissa tapahtuu useita ydinjakoja ennen sytoplasmista jakautumista.
Prosessi alkaa uran tai raon esiintymisestä, joka on merkitty metafaasisen plakin tasoon, varmistaen, että jako tapahtuu kromosomiryhmien välillä. Jälkipaikan merkitsee mitoottinen kara erityisesti, Ásteresin mikrotubulukset.
Merkityssä rakoissa on sarja mikrofilamentteja, jotka muodostavat renkaan, joka on suunnattu solukalvon sytoplasmiselle puolelle, koostuu suurelta osin aktiinista ja myosiinista. Nämä proteiinit ovat vuorovaikutuksessa keskenään, mikä mahdollistaa renkaan supistumisen uran ympärillä.
Tämä supistuminen syntyy näiden proteiinien filamenttien liukumalla vuorovaikutuksessa toistensa kanssa samalla tavalla kuin he tekevät esimerkiksi lihaskudoksissa.
Renkaan supistuminen syvenee aiheuttamalla "puristaminen" -vaikutus, joka lopulta jakaa emosolun, sallimalla tytärsolujen erottamisen sen sytoplasmisella sisällöllä kehityksessä.
Sytocinesis kasvisoluissa
Kasvien soluilla on soluseinä, joten niiden sytoplasminen jakoprosessi on erilainen kuin aiemmin kuvattu ja alkavat telfaasissa.
Uuden soluseinämän muodostuminen alkaa koota jäännöskaran mikrotubulukset, muodostuen framoplastille. Tämä lieriömäinen rakenne muodostuu kahdella mikrotubuluksen pelillä, jotka yhdistävät sen päissä ja joiden positiiviset navat upotetaan elektroniseen plakkiin päiväntasaajan tasoon.
Pienet vesikkelit Golgi -laitteesta, täynnä soluseinämän esiasteita, liikkuu framoplastien mikrotubulusten läpi päiväntasaajan alueelle, yhdistämällä solun plakin muodostamiseksi. Vesikulaisten sisältö on erotettu tälle levylle sen kasvaessa.
Tämä levy kasvaa, sulautuen plasmamembraaniin solun kehää pitkin. Tämä tapahtuu framoplastien mikrotubulusten jatkuvasta uudelleenorganisoinnista levyn reuna -alueella, jolloin enemmän rakkuloita voi liikkua tätä tasoa ja tyhjentää sen sisältöä.
Tällä tavoin tapahtuu tytärsolujen sytoplasminen erottaminen. Lopuksi sen sisällä olevan selluloosan mikrokuitujen vieressä olevan solulevyn pitoisuus mahdollistaa uuden soluseinämän muodostumisen loppuun.
Funktiot
Mitoosi on jakautumismekanismi soluissa, ja se on osa yhtä solusyklin faaseja eukaryooteissa. Yksinkertaisesti, voimme sanoa, että tämän prosessin päätehtävä on solun lisääntyminen kahdessa tytärsoluissa.
Yksisoluisilla organismeissa solujen jakautuminen tarkoittaa uusien yksilöiden tuottamista, kun taas monisoluisten organismien osalta tämä prosessi on osa täydellisen kehon kasvua ja oikeaa toimintaa (solunjako tuottaa kudosten kehitystä ja rakenteiden ylläpitämistä).
Mytisiprosessi aktivoidaan organismin vaatimusten mukaisesti. Esimerkiksi nisäkkäissä punasolut (punasolut) alkavat jakaa enemmän soluja, kun keho tarvitsee paremman hapen sieppauksen. Samoin valkosolut (leukosyytit) lisääntyvät, kun tartunnan on tarpeen torjua infektiota.
Sitä vastoin joistakin erikoistuneista eläinsoluista puuttuu käytännössä mitoosiprosessi tai on hyvin hidas. Esimerkki tästä on hermosoluja ja lihassoluja).
Yleensä ne ovat soluja, jotka ovat osa organismin sidekudosta ja rakenteellista kudosta ja joiden lisääntyminen on välttämätöntä vain silloin, kun solulla on jonkin verran virhettä tai heikkenemistä ja se on vaihdettava.
Kasvun säätely ja solujen jako.
Solujen kasvu- ja solujen jakautumisjärjestelmä on paljon monimutkaisempi monisoluisissa organismeissa kuin yksisoluisissa. Jälkimmäisessä resurssien saatavuus rajoittaa jälkimmäistä jäljennöstä.
Eläinsoluissa jako pidätetään, kunnes on positiivinen signaali, joka aktivoi tämän prosessin. Tämä aktivointi on kemiallisten signaalien muodossa naapurisoluista. Tämä mahdollistaa kudosten rajoittamattoman kasvun ja viallisten solujen lisääntymisen, mikä voi vakavasti vahingoittaa organismin elämää.
Voi palvella sinua: Peruskalvo: Ominaisuudet, rakenne ja toiminnotYksi mekanismeista, jotka kontrolloivat solujen kertomista, on apoptoosi, jossa solu kuolee (johtuen tiettyjen proteiinien tuotannosta, jotka aktivoivat itsensä tuhoamisen), jos sillä on huomattavia vaurioita tai sen tartunnan saaneet viruksella.
Solujen kehityksen säätely on myös estämällä kasvutekijöitä (kuten proteiini). Siten solut pysyvät rajapinnassa etenemättä solusyklin M -vaiheeseen.
Organismit, jotka toteuttavat sen
Mitoosiprosessi suoritetaan suurimmassa osassa eukaryoottisia soluja yksisoluisista organismeista, kuten hiivasta, jotka käyttävät sitä aseksuaalisena lisääntymisprosessina monimutkaisten monisoluisten organismien, kuten kasvit ja eläimet ja eläimet.
Vaikka solusykli on yleensä sama kaikille eukaryoottisoluille, yksisoluisten ja monisoluisten organismien välillä on merkittäviä eroja. Entisessä solujen kasvua ja jakautumista suosii luonnollinen valinta. Monisoluisissa organismeissa leviämistä rajoittavat tiukat kontrollimekanismit.
Yksisoluisissa organismeissa lisääntyminen tapahtuu kiihdytettynä, koska solusykli toimii jatkuvasti ja tytärsolut alkavat nopeasti kohti myitoosia jatkaakseen kyseistä sykliä. Kun taas monisoluisten organismien solut vievät huomattavasti enemmän aikaa kasvaa ja jakaa.
Kasvien ja eläinsolujen mitoottisten prosessien välillä on myös joitain eroja, kuten joissakin tämän prosessin vaiheissa, mutta periaatteessa mekanismi toimii samalla tavalla näissä organismeissa.
Solujen jako prokaryoottisissa soluissa
Prokaryoottinen solu Yleensä prokaryoottiset solut kasvavat ja jaetaan nopeammin kuin eukaryoottiset solut.
Organismeista, joilla on prokaryoottisia soluja (yleensä yksisoluiset tai joissakin monisoluisissa tapauksissa), puuttuu ydinkalvo, joka eristää geneettisen materiaalin ytimen sisällä, joten se on dispergoituna solussa, nukleoidia kutsulla alueella, jota kutsutaan nukleoidiksi. Näillä soluilla on tärkein pyöreä kromosomi.
Solujen jakautuminen näissä organismeissa on silloin paljon suorempaa kuin eukaryoottisoluissa, puuttuu kuvattu mekanismi (mitoosi). Niissä lisääntyminen suoritetaan binaarinen fissio -niminen prosessi, jossa DNA: n replikaatio alkaa tietyssä ympyräkromosomikohdassa (replikaation alkuperä tai ORIC).
Muodostuu kaksi alkuperää, jotka siirtyvät solun vastakkaisille puolille replikaation tapahtuessa, ja solu venyy, kunnes se saavuttaa kaksinkertaisen koon. Replikaation lopussa solukalvo kasvaa sytoplasmaan jakamalla progenitorisolun kahteen tyttäreen samalla geneettisellä materiaalilla.
Mitoosin kehitys
Eukaryoottisten solujen kehitys toi mukanaan genomin monimutkaisuuden lisääntymisen. Tämä merkitsi yksityiskohtaisempien jakautumismekanismien kehitystä.
Mikä edelsi mitoosia?
On hypoteeseja, jotka ehdottavat, että bakteerien jako on mitoosin edeltäjämekanismi. Binaariseen fissioon liittyvien proteiinien välillä on löydetty tietty suhde (joka voi olla ne, jotka ankkuroivat kromosomit tyttäreiden plasmamembraanin tiettyihin kohtiin) eukaryoottisten solujen tubuliinin ja aktiinin kanssa.
Jotkut tutkimukset osoittavat tietyt erityispiirteet nykyaikaisten yksisoluisten protistien jakautumisessa. Niissä ydinkalvo pysyy ennallaan mitoosin aikana. Replikoituneet kromosomit pysyvät ankkuroituneina tämän kalvon tiettyihin kohtiin, erottaen, kun ydin alkaa venyttää solun jakautumisen aikana.
Tämä osoittaa tietyn sattuman binaarisen fissioprosessin kanssa, jossa toistuneet kromosomit on kiinnitetty tiettyihin paikkoihin solukalvossa. Sitten hypoteesi ehdottaa, että protistit, jotka esittävät tämän laadun solujen jakautumisensa aikana.
Tällä hetkellä selityksiä siitä, miksi monisoluisten organismien eukaryoottisia soluja ei ole vielä kehitetty, on välttämätöntä, että ydinkalvo hajoaa solunjakoprosessin aikana.
Viitteet
- Albarracín, a., & Telulón,. -Lla. (1993). Soluteoria 1800 -luvulla. Akal Editions.
- Alberts, b., Johnson, a., Lewis, J., Raff, m., Roberth, k., & Walter, P. (2008). Solumolekyylin biologia. Garland Science, Taylor ja Francis Group.
- Campbell, n., & Reece, J. (2005). Biologia 7th Painos, AP.
- Griffiths, a. J -., Lewontin, R. C., Miller, J. H., & Suzuki, D. T. (1992). Johdanto geenianalyysiin. McGraw-Hill-Amerikanvälinen.
- Karp, g. (2009). Solu- ja molekyylibiologia: käsitteet ja kokeet. John Wiley & Sons.
- Loodish, h., Darnell, J. JA., Berk, a., Kaiser, c. -Lla., Krieger, m., Scott, M. P., & Matsudaira, P. (2008). Mollecular -solusbiologia. Macmillan.
- Segura-valdez, m. D -d. Lens., Cruz-Gómez, S. D -d. J -., López-Cruz, R., Zavala, G., & Jiménez-García, L. F. (2008). Mitoosin visualisointi atomivoimamikroskoopilla. Kärki. Erikoistunut aikakauslehti kemian biologisissa tieteissä, 11 (2), 87-90.