

Adenilato ciclasa -ominaisuudet, tyypit, toiminnot
- 4651
- 125
- Gustavo Runte DVM
Se Syklasa adenylaatti jompikumpi adenyylikilaasi ATP-muunnosta vastaava entsyymi, korkeaenergiamolekyyli, syklisessä AMP: ssä, tärkeässä signalointimolekyylissä, joka aktivoi erilaisia syklisistä riippuvaisia AMP-proteiineja, joilla on tärkeät fysiologiset toiminnot.
Sen aktiivisuutta säätelevät erilaiset tekijät, kuten hormonien, välittäjäaineiden ja muiden luonteeltaan muiden säätelymolekyylien (kaksiarvoiset kalsiumioonit ja G -proteiinit.
Cyclasa adenylaattikaavio (lähde: Käyttäjä Bensaccount.Wikipedia [julkinen alue] Wikimedia Commonsin kautta) Tämän entsyymin tärkein merkitys on katalysoidun reaktion, syklisen AMP: n, transsendenssissa, koska se osallistuu monien aineenvaihduntaan ja kehitykseen liittyvien soluilmiöiden hallintaan, samoin kuin vasteeseen eri ulkoisiin ärsykkeisiin.
Luonnossa sekä yksisoluiset organismit (suhteellisen yksinkertaiset) että suuret ja monimutkaiset monisoluiset eläimet käyttävät syklistä AMP: tä toisena lähettiläänä ja siten entsyymeihin, jotka tuottavat sitä.
Fylogeneettiset tutkimukset osoittavat, että nämä entsyymit ovat peräisin yhteisestä esi -isästä ennen eubakteerien ja eukaryootien erottamista, mikä viittaa siihen, että syklisellä AMP: llä oli erilaisia toimintoja, ehkä liittyvät ATP -tuotantoon.
Tällainen lausunto on mahdollista hyväksyä, koska adenilaattisyklaasin katalysoima reaktio on helposti palautettava, mikä voidaan havaita ATP: n synteesin tasapainossa vakiona (ATP: n synteesi (K -k -Eq ≈ 2.1 ± 0.2 10-9 M2-A.
[TOC]
Ominaisuudet ja rakenne
Suurin osa eukaryoottisista syklastien adenylaattientsyymeistä liittyy plasmamembraaniin, mutta bakteereissa ja nisäkkäiden siittiöissoluissa ne löytyvät liukoisina proteiineina sytosolissa.
Hiivoissa ja jotkut bakteerit ovat perifeerisiä membraaniproteiineja, kun taas tietyissä amoeba -lajeissa ne ovat molekyylejä, joilla.
Rakenteelliset ominaisuudet
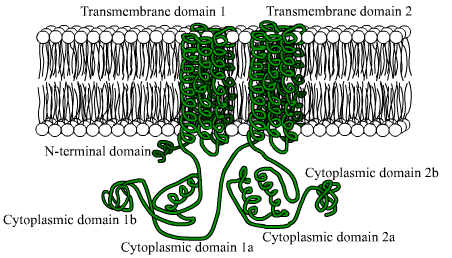
Ne ovat proteiineja, jotka koostuvat suurista polypeptidiketjuista (yli 1.000 aminohappojäte), jotka ylittävät plasmamembraanin 12 kertaa kahdella alueella, jotka koostuvat kuudesta kalvon läpäisevästä domeenista konformaation alfa.
Jokainen transmembraniaalinen alue erotetaan suurella sytosolisella domeenilla, joka vastaa katalyyttisestä aktiivisuudesta.
Voi palvella sinua: Afrikan kasvisto ja eläimistö: edustavat lajitEukaryoottisten organismien joukossa on joitain syitä, jotka on säilytetty näiden entsyymien amino -päätealueen fragmentissa, samoin kuin noin 40 kDa: n sytoplasmisen domeenin, jonka hydrofobiset osat ovat rajattu.
Katalyyttinen alue
Reaktio, jota nämä entsyymit katalysoivat, toisin sanoen ruokavalion sidoksen muodostuminen OH -ryhmän nukleofiilisen hyökkäyksen kautta 3 '-asennossa nukleosiditrfosfaatin fosfaattiryhmään asennossa 5', riippuu yhteisestä rakenteellisesta motiivista, joka tunnetaan nimellä The the the Verkkotunnuksen palmu ".
Tämä "Palman" alue koostuu syystä "βαβαβ"(" P ", joka haluaa merkitä β-panostettuja ja" α "-levyjä alfapotkurille) ja sillä on kaksi muuttumatonta vastaavaa happotähdettä, jotka koordinoivat katalyysistä vastaavaa metalli-ionia, jotka voivat olla kaksiarvoisia magnesium- tai magnesium-ionit sinkki.
Monet näiden entsyymien kvaternääriseen rakenteeseen liittyvät tutkimukset ovat paljastaneet, että niiden katalyyttinen yhtenäisyys on dimeerinä, jonka muodostuminen riippuu läpäisevästä segmentistä, jotka on kerätty endoplasmisessa retikulumissa proteiinin muodostumisen aikana.
Sijainti
On määritetty, että kuten monet kalvon kattavat proteiinit, kuten g -proteiinit, ne, joilla on fosfatidyylinositoli -ankkureita ja monia muita, adenyylisääkkeitä löytyy alueilta tai erityisiltä membraanin mikrodomeeneihin, jotka tunnetaan nimellä “lipidibalsas” (englanniksi ”(englanniksi” (englanniksi ”(englanniksi” "Lipidilautta"-A.
Näissä kalvodomeeneissa voi olla jopa satoja nanometrejä, ja ne koostuvat pääasiassa kolesterolista ja spingolipideistä, joissa on pääosin tyydyttyneitä rasvahappoketjuja ja suuria pitkiä pituisia, mikä aiheuttaa niille vähemmän juoksevuutta ja mahdollistaa transmembraal -segmenttien sopeutumisen erilaisten proteiinien mukautumiseen, ja ne voivat sopeutua transmembraalisiin segmentteihin eri proteiineja.
Cyclasas -adenilaatti on myös löytänyt olevansa liitetty lipidilauttojen osa -alueisiin, jotka tunnetaan nimellä "Caveolas" (englanti "(englanti"Caveolae "), jotka ovat pikemminkin kolesterolin runsaaseen membraanin ja siihen liittyvää proteiinia, jota kutsutaan caveolinaksi.
Kaverit
Luonnossa on kolmen tyyppisiä hyvin määriteltyjä syklakentteja adenylaattia ja kaksi, jotka ovat nykyisiä keskustelua.
Voi palvella sinua: Oxihemoglobin: Ominaisuudet, rakenne ja unionin käyrä- Luokka I: Niitä on läsnä monissa gram -negatiivisissa bakteereissa, kuten JA. koli, Esimerkiksi, missä reaktion syklinen AMP -tuotteella on toimintoja, kuten ligandi transkriptiotekijöille, jotka vastaavat katabolisten operaatioiden säätelystä.
- Luokka II: Niitä löytyy joistakin bakteerigenrejen patogeeneistä, kuten Basilli jompikumpi Bordetella, missä ne toimivat solunulkoisena toksiinina. Ne ovat proteiineja, joita isäntäkalvoduliini aktivoi (bakteereissa).
- Luokka III: Ne tunnetaan nimellä "universaali" luokka ja ne ovat fylogeneettisesti suhteessa Guanilato Cyclasasiin, jotka täyttävät samanlaiset toiminnot. Niitä löytyy sekä prokaryooteista että eukaryotaista, joissa niitä säätelevät eri teillä.
Adenylaatti nisäkkäiden syklasas
Nisäkkäissä ainakin yhdeksän näitä entsyymejä on kloonattu ja kuvattu.
Näillä on monimutkaiset rakenteet ja kalvotopologiat, samoin kuin kaksois katalyyttiset domeenit, jotka ovat ominaisia.
Nisäkkäiden osalta isoformeihin viitattu nimikkeistö vastaa vaihtovirheitä (kirjoittanut Syklasa adenylaatti) ja numeroon 1 - 9 (AC1 - AC9). Myös AC8 -entsyymin kaksi varianttia on raportoitu.
Näissä eläimissä olevat isomuodot ovat vastineita niiden katalyyttisten kohtien primaarirakenteen ja kolmen dimensioisen rakenteen sekvenssin suhteen. Yhden näiden entsyymien sisällyttäminen kussakin "tyypissä" liittyy pääasiassa kunkin isomuodon säätelymekanismeihin.
Heillä on ekspressiokuvioita, jotka ovat usein kudosspesifisiä. Aivoista löydät kaikki isoformit, vaikka jotkut ovat rajoitettuja keskushermoston tietyille alueille.
Funktiot
The main function of the enzymes belonging to the Cylasa Adenylate family is to transform the ATP into cyclic AMP and for this they catalyze the formation of an intramolecular dieter link 3 ' - 5' (reaction similar to that catalyzed by the DNA polymerases), With Pyrofosfaattimolekyylin vapautuminen.
Nisäkkäissä, jotka voidaan saavuttaa erilaiset variantit, on liitetty solujen lisääntymiseen, etanoliriippuvuuteen, synaptiseen plastisuuteen, lääkkeen riippuvuuteen, vuorokausipäiväiseen rytmiin, hajujen stimulaatioon, oppimiseen ja muistiin.
Voi palvella sinua: lois: Konsepti, tyypit ja esimerkitJotkut kirjoittajat ovat todenneet, että syklasas -adenylaatissa voi olla lisätoiminto, kuten kuljetusmolekyylit tai, mikä on sama, kanavaproteiinit ja ioniset kuljettimet.
Nämä hypoteesit on kuitenkin todistettu vain näiden entsyymien transmbranal -segmenttien järjestelystä tai topologiasta riippuen, jolla on joitain rakenteellisia homologioita tai samankaltaisuuksia (mutta ei sekvenssiä) tietyillä ionin kuljetuskanavilla.
Sekä syklinen AMP että PPI (pyrofosfaatti), jotka ovat reaktion tuotteita, on toiminnot solutasolla; Mutta sen merkitys riippuu organismista missä he ovat.
Säätö
Adenyylisilasien välinen suuri rakenteellinen monimuotoisuus osoittaa suuren alttiuden monille säätelymuodoille, mikä antaa niiden integroida moniin solujen signalointireiteihin.
Joidenkin näiden entsyymien katalyyttinen aktiivisuus riippuu alfaketoasideista, kun taas toisilla on paljon monimutkaisempia säätelymekanismeja, joihin liittyy säätely -alayksiköitä (stimulaatiolla tai estämällä), jotka riippuvat esimerkiksi kalsiumista ja muista tekijöistä, sekä muut, samoin kuin muut, samoin kuin muihin proteiinit.
Joidenkin G -proteiinien alayksiköitä säätelevät monia syklasasia adenilaattia (estävät niiden toimintaa), kun taas toiset käyttävät pikemminkin aktivaattoreita.
Viitteet
- Cooper, D. M. F. (2003). Adenylyylisyklasien ja leirin sääntely ja organisointi. Lehden biokemiallinen, 375, 517-529.
- Cooper, D., Mons, n., & Karpen, J. (tuhatyhdeksänsataayhdeksänkymmentäviisi). Adenylyylisyklaasit ja kalsiumin ja cAMP Sigalingin vuorovaikutus. Luonto, 374, 421-424.
- Danchin, a. (1993). Adenylyylisyklasien fylogeny. Edistyminen toisen lähettilään ja fosfoproteiinitutkimuksessa, 27, 109-135.
- Hanoune, J., & Lykkää, n. (2001). Adenylyylisyklaasin isoformien säätely ja rooli. Lupa. Rev. Farmakoli. Toksikoli., 41, 145-174.
- Raja, u., & Schultz, J. JA. (2003). Luokan III adenylyylisyklasi: monikäyttöinen signalointimoduulit. Solujen signalointi, viisitoista, 1081-1089.
- Tang, W., & Gilman,. G. (1992). Adenylyylisyklasit. Solu, 70, 669-672.
- « Äänen diffraktio, josta se koostuu, esimerkkejä, sovelluksia
- Santa Alliance -tausta, tavoitteet, seuraukset »