

Bilapa -lipidiominaisuudet, rakenne, toiminnot

- 4900
- 265
- Gabriel Fahey
Se Lipidikerroksinen Se on ohut laminaarikalvo, bimolekulaarinen, amfipaattiset lipidit, ts. Ne sisältävät hydrofobisen osan ja toisen hydrofiilisen osan. Sillä on erittäin matala läpäisevyys ioneille, samoin kuin useimpien vesiliukoisten molekyylien suhteen, mutta se on hyvin läpäisevä vettä.
Vesipitoisissa liuoksissa polaariset lipidit, kuten fosfoglyseridi. Näissä rakenteissa hydrofiiliset polaaristen lipidien päät ohjataan ulkoisesti ottamaan yhteyttä veteen, kun taas hännät (hydrofobiset) on järjestetty vastakkaiseen päähän.
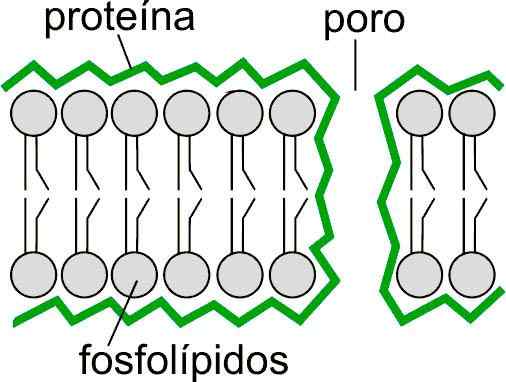
Kaavio mahdollisista lipidijärjestelyistä huokosen reunalla lipidikerroksen läpi. Otettu ja muokattu osoitteesta: mdougm [julkinen alue]. Elävien olentojen solukalvot muodostavat pääasiassa fosfolipidit ja glykolipidit, jotka muodostavat lipidikerroksen. Tämä kaksikerroksinen on läpäisevyyseste, joka mahdollistaa suolojen ja soluelektrolyyttien sisäisen sisällön säätelyn. Tämän saavuttamiseksi heillä on rakenteita, joita kutsutaan ionipumppuiksi.
Ensimmäiset tutkijat, jotka ehdottivat lipidi -bilap -mallia solukalvoille, olivat lääkärit Evert Gorter ja F. Grendel (1925), Leidenin yliopistosta, Holland, malli, joka vahvistettiin vuonna 1950 elektronisten mikroskopian avulla.
Lipidikiskojen nykyisiä ja mahdollisia käyttötarkoituksia on useita, mutta tähän mennessä menestynein kaupallisesti puhuminen on ollut keinotekoisten vesikkelien (liposomien) käyttö lääkkeiden antamiseen syöpäpotilaille syöpäpotilaille.
[TOC]
Ominaisuudet
Lipidikerrokset ovat laminaarisia, erittäin ohuita ja hauraita rakenteita, joilla on joitain biologisesti tärkeitä ominaisuuksia, kuten:
Läpäisevyys
Yksi lipidikerroksen pääominaisuuksista on sen selektiivinen läpäisevyys. Nämä kalvot ovat todellakin hyvin vedenpitäviä ioneille ja useimmille polaarimolekyyleille, vesi on tärkeä poikkeus, koska se voi helposti ylittää kalvon.
Esimerkki tästä selektiivisestä läpäisevyydestä on natrium ja kalium, joiden ionit kulkevat kalvon läpi yli miljoona kertaa hitaammin kuin vesi. Toisaalta indoli, heterosyklinen orgaaninen yhdiste, ylittää kalvon nopeudella tuhat kertaa korkeampi kuin tryptofaani, toinen tämän rakenteen kaltainen molekyyli.
Jo ennen kalvon kaksinkertaista luonnetta tiedossa, tiedemies Charles Overton kertoi (1901), että koon molekyylien läpäisevyyskertoimet liittyvät suoraan orgaanisissa liuottimissa ja vedessä esiintyvään suhteelliseen liukoisuuteen ja vedessä.
Epäsymmetria
Jokainen kalvon muodostava kerros on rakenteellisesti ja toiminnallisesti erilainen kuin muut. Funktionaalinen esimerkki tästä epäsymmetriasta on natriumkaliumpumppu. Tätä pumppua on läsnä valtaosan suurimpien organismien solujen plasmamembraanissa.
Naa pommi+ - K -k -+ Se on suunnattu siten, että se karkottaa Na+ solun sisällä, ottaen käyttöön k -ionit+. Lisäksi tämä kuljetusväline tarvitsee energiaa ATP: n muodossa aktivointia varten ja sitä voidaan käyttää vain, jos se on solun sisällä.
Kunkin kerroksen komponentit ovat myös erilaisia, membraaniproteiinit syntetisoidaan ja asetetaan epäsymmetrisesti glukolipidien kahteen.
Voi palvella sinua: Crocodylus acutusEsimerkiksi punasolujen tapauksessa sphylingomyeline ja fosfatidyylikoliinit sijaitsevat kalvon ulkokerroksessa, kun taas fosfatidyleletalamiini ja fosfatidyyliseriini ovat sisäisiä. Kolesteroli on kuitenkin ainesosa, molemmat kerrokset.
Yksi epäsymmetrian syistä fosfolipidien jakautumisessa on, että suurin osa näistä aineosista syntetisoidaan solussa ja siksi ne sisällytetään alun perin sisäkerrokseen, ja siten jotkut niistä siirtyvät ulkokerrokseen joidenkin nimettyjen entsyymien avulla nimeltään flipasas.
Sujuvuus
Lipidikynät eivät ole jäykkiä rakenteita, mutta päinvastoin, ne ovat nestemäisiä ja dynaamisia rakenteita, joissa lipidit ja monet proteiinit ovat moraalisesti moraalisesti moraalisesti moraalisesti.
Lipidit leviävät sivusuunnassa kalvoon keskimääräisellä nopeudella 2 um sekunnissa. Proteiinien sivuttaissiirto kaksikerroksissa voi silti vaihdella proteiinityypistä riippuen; Vaikka jotkut ovat yhtä nopeasti kuin lipidit, toiset käytännössä pysyvät liikkumattomina.
Poikittainen diffuusio, jota kutsutaan myös flip-flopiksi, on toisaalta paljon hitaampi lipideille, eikä sitä ole koskaan havaittu proteiineissa.
Toisaalta kalvon juoksevuus voi vaihdella lipidirasvahappojen suhteellisesta järjestyksestä riippuen. Kun kaikki rasvahapot on tilattu, Bilay.
Nämä muutokset voivat johtua lämpötilan vaihteluista; Siirtyminen kiinteästä nestetilaan tapahtuu äkillisesti, kun lämpötila ylittää sulamislämpötilan nimellä olevan kynnyksen, joka riippuu rasvahappoketjujen pituudesta, samoin kuin sen tyydyttymättömyyden asteesta.
Membraanin muodostavat lipidit ovat luonteeltaan erilaisia, ja siksi ne voivat esiintyä erilaisia sulamislämpötiloja. Tämän vuoksi eri lämpötiloissa ne voivat esiintyä samanaikaisesti kiinteissä ja nestemäisissä vaiheissa samassa kaksikerroksessa.
Muut ominaisuudet
Lipidikerroksisilla, kovalenttisten vuorovaikutusten ja van der Waalsin houkuttelevien voimien ansiosta on taipumus olla laaja, samoin kuin itsessään, jotta ei ole äärimmäisiä alttiita ääripäitä. Sen kyky itsepalata on myös ominainen, koska jatkuvuuden puute ei ole energisesti suotuisa rakenteelle.
Rakenne
Lipidikaksoiskerroksen rakenteen selittämiseksi on erilaisia malleja:
Davson ja Danilli -malli
Sitä ehdotettiin vuonna 1935, ja väittää, että kalvot sisältävät jatkuvan hiilivetyvaihetta, joka tarjoaa kalvon muodostavat lipidit.
 Davson ja Danielli -solukalvomalli. Otettu ja muokattu osoitteesta: Miguelferig [julkinen alue].
Davson ja Danielli -solukalvomalli. Otettu ja muokattu osoitteesta: Miguelferig [julkinen alue]. Yhtenäinen kalvomalli
J: t.D -d. Robertson, tämä hypoteesi on Davson- ja Danilli -mallin muutos. Hän postuloi, että yksikkökalvo koostui kaksoiskerroksesta sekoitettuja polaarisia lipidejä.
Nämä lipidit suuntautuivat hiilivetyketjuilla sisäänpäin, muodostaen jatkuvan hiilivetykerroksen, kun taas hydrofiiliset päät osoittivat vastakkaiseen suuntaan.
Lisäksi tämä yhtenäinen membraani peitettiin molemmilla puolilla yksinkertaisella proteiinimolekyylikerroksella, joka oli järjestetty.
Globaali malli
Tunnetaan myös nimellä Subunites -malli. Tämän mallin mukaan kalvot muodostuisivat lipoproteiinien toistuvien alayksiköiden mosaiikilla välillä 4,0 - 9,0 nm.
Voi palvella sinua: Hammer Head Bat: Ominaisuudet, elinympäristö, lisääntyminen, ruokaNeste mosaiikkimalli
Ehdotti S.J -. Laulaja ja g.Lens. Nicholson vuonna 1972 ja on hyväksytty malli. Sen mukaan membraanifosfolipidit järjestetään kaksoiskerroksina muodostaen nestekiteitä matriisin.
Tämän mallin mukaan yksittäiset lipidimolekyylit voivat matkustaa vapaasti.
Proteiinien, jotka ovat osa kaksikerroksista, mallin mukaan on oltava pallomaisia. Lisäksi jotkut proteiinit upotetaan osittain kaksikerrokseen, kun taas toiset olisivat täysin upotettuja tähän.
Globaalien proteiinien tunkeutumisaste Bilayssa.
Sävellys
Luonnolliset kaksikerrokset koostuvat pääasiassa fosfolipideistä. Nämä ovat glyserolista peräisin olevia yhdisteitä, joille on tunnusomaista hydrofiilinen pää ja kaksi hydrofobista pyrstöä.
Kun fosfolipidit joutuvat kosketuksiin veden kanssa, voidaan järjestää eri tavoin. Vakain tapa on kuin kaksikerroksinen, jonka hännät suuntautuvat sisämaahan ja päätä kaksikerroksen ulkopuolelle.
Glykolipidit ovat myös osa lipidikerroksista. Nämä yhdisteet, kuten nimestä päätetään.
Toinen tärkeä kalvon aineosa on kolesteroli, skaponitoimaton lipidi. Se on läsnä sekä kaksikerroksen sisä- että ulkokerroksessa. Se on runsaammin plasmamembraanissa kuin organelien kalvossa.
Kalvoilla on myös monenlaisia proteiineja, jotka voivat olla kahden tyyppisiä, ulkoisia tai sisäisiä. Ulkopuoliset tai perifeeriset proteiinit liittyvät kalvon huonoihin ammattiliittoihin ja voivat helposti erottaa ne niistä.
Luonnolliset tai kiinteät proteiinit liittyvät voimakkaasti Bilayiin. Ne edustavat noin 70% kalvoproteiineista. Joillakin heistä on signaalien funktio solun ulkopuolelta ja sen lähetys sisällä.
Muut proteiinit liittyvät kahden erilaisen kaksikerroksen fuusioon. Heidän joukossaan ovat ne, jotka sallivat siittiöiden liiton ovuulen kanssa hedelmöityksen aikana; myös ne, jotka sallivat virusten tunkeutumisen isäntäsoluihin.
Lisäksi ionipumput ovat kattavia proteiineja, jotka ylittävät Bilayn.
Funktiot
Lipidi -bilayn tärkein biologinen toiminta. Ilman tätä osastojen välistä fyysistä rajaamista, elämä olisi mahdotonta, koska tiedämme sen.
Tämä toiminto on niin tärkeä. Poikkeusta edustaa jotkut kaarilajit, joissa kalvo on lipidi -yksikerros.
Lipidikerrokset osallistuvat interneuronaalisen hermon impulssin siirtoon. Neuroneja ei ole fyysisesti kytketty toisiinsa, vaan erotettuna lyhyellä tilalla, nimeltään Sinapsis. Tämän interneuronaalisen tilan tallentamiseksi on kyse välittäjäaineiden kanssa ladattujen vesikkelien kanssa.
Voi palvella sinua: eläimet, jotka hengittävät stomatallaToinen kaksikerroksinen toiminta on toimia rakenteellisena perustana tai tukemaan luurankoa, joihin jotkut kuljetusjärjestelmät ovat olleet voimakkaasti yhtenäisiä ja joitain entsyymejä.
Organelit, joissa on lipidikerroksinen
Prokaryooteissa lipidi bilay.
-Organelit, joissa on kaksi lipidikaksoiskerrointa
Ydin
Eukaryoottisissa soluissa esiintyvä solu organeli ja joka sisältää suurimman osan kromosomeissa järjestetystä geneettisestä materiaalista.
Ydinmembraani muodostuu kahdella lipidillä, jotka on erotettu tilalla, jota kutsutaan perinukleaariksi. Molempia kerroksia kutsutaan ulkoiseksi ja sisäiseksi ydinmembraaniksi, ja ne erotetaan sen proteiinikoostumuksella.
Mitokondria
Organo, joka on vastuussa solujen hengityksestä, jota toimittaa solun aktiivisuuteen tarvittava energia. Siinä on kaksoiskalvo, sileä ulkoinen ja taitettu sisäinen muodostettava laminaarinen tai digitiformi harja.
Tällaisten laskosten tehtävänä on lisätä sisäpinta -alaa, mikä on paikka, jossa metaboliset reaktiot tapahtuvat.
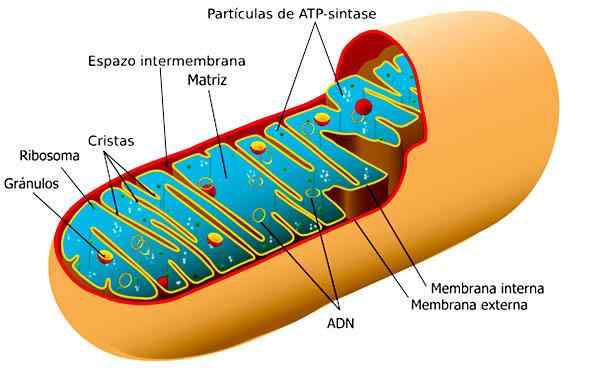 Mitokondria. Otettu ja muokattu osoitteesta: Ladyofhats [CC0].
Mitokondria. Otettu ja muokattu osoitteesta: Ladyofhats [CC0]. Kloroplasti
Organelle, joka on läsnä ylemmissä kasveissa ja muissa fotoautotrofisissa eukaryoottisissa organismeissa. Se esittelee kaksi samankeskistä lipidiä, jotka on erotettu intermembranoisella tilalla. Ulompi kerros on huokoisempi kuin sisäinen johtuen proteiineista, joita kutsutaan poriiniksi.
-Organelit, joissa on lipidikerroksinen
Plasmamembraanin lisäksi, jota on puhuttu laajasti tässä artikkelissa, muut organelit, kuten endoplasminen retikulum, Golgi -laitteisto ja lysosomit ovat yksi lipidikerroksinen.
Endoplasminen retikulum (uudelleen)
Liittyvät sytoplasman membraanikompleksit (Re -regged) tai ei (sileä uudelleen) ribosomeihin ja jotka osallistuvat lipidien ja fosfolipidien (sileä RE) tai peptidien ja proteiinien (RE Ruguure) synteesiin niiden seinämiin kiinnitettyjen ribosomien ansio.
Golgin laite
Proteiiniaineiden varastointiin, modifiointiin ja pakkaamiseen osallistuvien sileiden seinäkalvojen kompleksi.
Lysosoomat
Vesikulaariset organelit, jotka sisältävät entsyymejä, jotka osallistuvat vieraiden materiaalien hajoamiseen. Ne myös hajottavat ei -tarpeettomat solukomponentit ja jopa vaurioituneet tai kuolleet solut.
Sovellukset
Lipidikalapöydän pääsovellus sijaitsee lääketieteen alalla. Lipidi -kaksikerroksisten rajatut vesikulaariset rakenteet ovat vesikulaarisia rakenteita. Ne muodostuvat keinotekoisesti fosfoglyseridien vesipitoisista suspensioista.
Jos ionit tai molekyylit sisältyvät vesipitoiseen suspensioon, jotkut näistä elementeistä sisältävät liposomien sisällä. Näiden periaatteiden perusteella lääkkeet on kapseloitu liuokseen liposomeihin.
Lääkkeiden tyytyväiset liposomit toimitetaan potilaalle. Kun se on sisätiloissa. Määränpäässä heidän sisällönsä on rikki ja julkaistu.
Lipidi -bilapasten, kuten biosensorien, käyttö sairauksien diagnosoinnissa on myös testattu; samoin kuin biologisten aseiden mahdollinen havaitseminen. Lopuksi se on testattu onnistuneesti lääkkeen läpäisevyyden määrittämiseksi.
Viitteet
- JOMPIKUMPI.S. Andersen, II Koeppe, ja. Roger (2007). Kaksikerroksen paksuus ja membraaniproteiinifunktio: ANE Energinen näkökulma. Biofysiikan ja biomolekyylisen rakenteen vuosittainen katsaus.
- Lipidikerroksinen. Ecuured. Ecroved.com.
- Lipidikerroksinen. Wikipediassa. Toipunut Wikipediasta.org.
- -Lla. Lehninger (1978). Biokemia. Omega Editions, S.-Lla.
- Lens. Stryer (1995). Biokemia. W -.H. Freeman and Company, New York.
- R -.B -. Gennis (1989). Biomembraani. Springer-Verlag.
- M.S. Bretscher (1972). Biologisten kalvojen epäsymmetrinen lipidikerroksen rakenne. Luonto uusi biologia.
- « Zenaida -asaattiset ominaisuudet, elinympäristö, ruoka
- Harmonia Axyridis -ominaisuudet, elinkaari, elinympäristö »