

Cystring -luokittelu, koulutus, kehittäminen ja segmentointi
- 1688
- 220
- Eddie Hackett
Hän tsygootti Se määritellään soluksi, joka johtuu kahden sukusolun, yhden naisen ja yhden maskuliinisen välisen fuusion perusteella. Geneettisen kuorman mukaan zygootti on diploidi, mikä tarkoittaa, että se sisältää kyseisen lajin täydellisen geneettisen kuorman. Tämä johtuu siitä, että siitä peräisin olevat sukusolut sisältävät puolet lajien kromosomeista.
Se tunnetaan usein munaksi ja koostuu rakenteellisesti kahdesta pronudeista, jotka tulevat kahdesta siitä peräisin olevasta sukusolusta. Samoin sitä ympäröi pelukidinen alue, joka täyttää kolminkertaisen funktion: Vältä jonkin muun siittiöiden välillä, pidä solut, jotka johtuvat zygootin ensimmäisistä jakoista yhdessä ja estävät toteutuksen tapahtuvan, kunnes tsygootti saavuttaa alueen ihanteellisen. kohtu.
Zygootin kehitys. Lähde: CNX OpenStax [CC BY 4.0 (https: // creativecommons.Org/lisenssit/by/4.0)] Zygootin sytoplasma, samoin kuin siihen sisältyvät organelit, ovat äidin alkuperää, koska ne tulevat munasolusta.
[TOC]
Luokittelu
Zygootti luokitellaan kahden kriteerin mukaan: Vienteuksen määrä ja vitallus -organisaatio.
-Zygoottityypit Vitelon määrän mukaan
Sygootin vitalluksen määrästä riippuen tämä voi olla:
Oligolesito
Yleensä oligolesito -zygote on sellainen, joka sisältää hyvin vähän Viteloa. Samoin useimmissa tapauksissa ne ovat pienentyneet kokoa ja ytimellä on keskeinen sijainti.
Utelias tosiasia on tämän tyyppinen muna, ne ovat lähinnä toukkia, joilla on vapaa elämä.
Eläintyyppiä, joissa tämän tyyppistä tsygoottia arvostetaan, ovat piikien, kuten siilit ja meritähdet; Jotkut madot, kuten Plainmint ja nematodit; Nilviäiset, kuten etanot ja mustekalat; ja nisäkkäät ihmisenä.
Mesolekki
Tämä on sana, joka koostuu kahdesta sanasta, "meso", joka tarkoittaa puolta ja "lecito", joka tarkoittaa Viteloa. Siksi tämäntyyppinen tsygootti on sellainen, jolla on kohtalainen määrä villuusta. Samoin tämä sijaitsee pääasiassa yhdellä zygote -napoista.
Tämäntyyppinen muna edustaa joitain selkärankaisia, kuten sammakkoeläimiä, joita edustavat sammakot, rupikonnat ja salamanderit, muun muassa.
Polilecito
Sana polyilecito muodostuu sanoilla "Poli", mikä tarkoittaa paljon tai runsaasti, ja "lecito", mikä tarkoittaa Viteloa. Tässä mielessä Polilecito Zygote on sellainen, joka sisältää suuren määrän Viteliä. Tämän tyyppisessä zygootissa ydin on Vitle -alueen keskeisessä asennossa.
Polilecito -zygootti on tyypillinen lintuille, matelijoille ja joillekin kaloille, kuten haille.
Zygote -tyypit Vitellus -organisaation mukaan
Vienteuksen jakautumisen ja organisaation mukaan zygootti luokitellaan seuraavasti:
Isolekki
Sana isolecito koostuu "ISO", mikä tarkoittaa samaa ja "lecito", mikä tarkoittaa Viteloa. Sillä tavalla, että isolsitotyyppi zygootti on sellainen, jossa Vientelli esittelee homogeenisen jakautumisen koko käytettävissä olevaan tilaan.
Tämäntyyppinen zygootti on tyypillinen eläimille, kuten nisäkkäille ja merisiileille.
Telolekyyli
Tämän tyyppisessä zygootissa vill on runsaasti ja miehittää melkein kaikki käytettävissä olevat tilat. Sytoplasma on melko pieni ja sisältää ytimen.
Tämä zygote edustaa kaloja, lintuja ja matelijoita.
Centroletes
Kuten nimellä päätetään, tämän tyyppisissä munissa Vitle on keskeisessä asennossa. Samoin ydin sijaitsee Vientisen keskustassa. Tälle zygotille on ominaista soikea muoto.
Tämäntyyppinen zygootti on tyypillinen niveljalkaisten ryhmän jäsenille, kuten arachnidit ja hyönteiset.
Zygootin muodostuminen
Zygootti on solu, joka muodostuu heti hedelmöitysprosessin jälkeen.
Hedelmöitys
Hedelmöitys on prosessi, jolla naispuoliset ja miespuoliset sukusolut liittyvät. Ihmisillä naaras -zygootti tunnetaan nimellä Ovule ja uros zygote kutsutaan siittiöiksi.
Se voi palvella sinua: bioelementit: luokittelu (ensisijainen ja toissijainen)Samoin hedelmöitys ei ole yksinkertainen ja yksinkertainen prosessi, mutta se koostuu sarjasta vaiheita, jokainen erittäin tärkeä, nimittäin:
Kosketus ja tunkeutuminen säteilyssä kruunussa
Kun siittiöt luovat ensimmäisen kontaktin munasolun kanssa, se tekee niin SO: n nimellä Pelukide -alueella. Tämä ensimmäinen kontakti on transsendenttinen merkitys, koska se palvelee jokaiselle suomalaista tunnistaakseen toisen, määrittäen, kuuluvatko ne samoihin lajeihin.
Samoin tässä vaiheessa siittiö pystyy ylittämään munasolun ympäröivät solukerroksen ja että kokonaisuutena tunnetaan säteilevänä kruununa.
Pystyäkseen ylittämään kyseisen solukerroksen, siittiöt erittää entsymaattisen aineen, nimeltään hyaluronidaasi, joka auttaa sinua prosessissa. Toinen elementti, joka antaa siittiöiden tunkeutua tähän munasolun ulkokerrokseen, on hännän kiihkeä liike.
Johdanto Pelucidan alueelle
Kun siittiöt ovat ylittäneet säteilyn kruunun, siittiöt kohtaavat toisen esteen munasolun tunkeutumiselle: pelukidialue. Tämä ei ole muuta kuin munasolua ympäröivä ulkokerros. Se koostuu pääasiassa glykoproteiineista.
Kun siittiöpää joutuu kosketuksiin pelukidialueen kanssa, acrosomisen reaktion niminen reaktio laukaisee. Tämä koostuu siittiöiden vapautumisesta entsyymeistä, jotka tunnetaan spermiolisiinina kokonaisuutena. Nämä entsyymit varastoidaan acrosoma -nimisen siittiöiden pään tilaan.
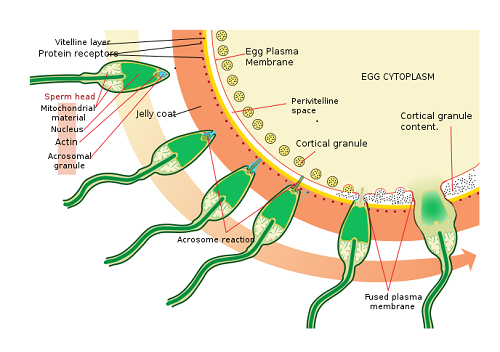 Akrosominen reaktio. Lähde: Ladyofhats. [Julkinen verkkotunnus]
Akrosominen reaktio. Lähde: Ladyofhats. [Julkinen verkkotunnus] Spermiolisiinit ovat hydrolyyttisiä entsyymejä, joiden päätehtävänä on pelukidialueen hajoaminen, jotta ne lopulta tunkeutuvat munasoluun.
Kun akrosominen reaktio alkaa, siittiöissä käynnistetään myös sarja rakenteellisia muutoksia sen kalvon tasolla, jonka avulla voit yhdistää kalvosi ovuulen kanssa.
Kalvofuusio
Seuraava vaihe hedelmöitysprosessissa on kahden sukusolun kalvojen fuusio, ts. Ovule ja siittiöt.
Tämän prosessin aikana munasarjassa tapahtuu sarjaa muunnos.
Ensinnäkin muodostuu lannoituskartio, jonka kautta siittiö- ja munasolujen kalvot joutuvat suoraan kosketukseen, jotka päättyvät yhdistämällä.
Samanaikaisesti tähän, munasolujen kalvotasolla ionin mobilisaatio, kuten kalsiumia (CA+2), Vety (H+) ja natrium (na+), joka tuottaa kalvon niin kutsutun depolarisaation. Tämä tarkoittaa, että normaalisti on napaisuus.
Samoin munasolun membraanin alla on rakenteita, joita kutsutaan aivokuoren rakeiksi, jotka vapauttavat niiden sisällön munasolun ympäröivään tilaan. Tämän avulla saavutetaan estämään siittiöiden tarttuminen munasolusta, joten he eivät voi lähestyä tätä.
Ovule -ytimien ja siittiöiden fuusio
Lopuksi zygootin muodostamiseksi on välttämätöntä, että siittiöt ja munasolut liittyvät.
On syytä muistaa, että sukusolut sisältävät vain puolet lajien kromosomien lukumäärästä. Ihmisen tapauksessa se on 23 kromosomia; Siksi kaksi ytimtä on yhdistettävä diploidisolujen muodostamiseksi, lajin täydellinen geneettinen kuormitus.
Kun siittiö saapuu munasoluun, DNA sisältää kaksinkertaistuneen, samoin kuin munasolun dna. Sitten molemmat pronucelot löytyvät toisen vierekkäin.
Välittömästi membraanit, jotka erottavat sekä hajoavat, että tällä tavalla kussakin olevat kromosomit voivat kokoontua heidän vastineensa kanssa.
Mutta kaikki ei lopu täältä. Kromosomit sijaitsevat solun (zygootin) päiväntasaajan navalla aloittamiseksi monista monista mitoottisista jakoista segmentointiprosessissa.
Voi palvella sinua: apolipoproteiini E: Ominaisuudet, toiminnot, sairaudetZygootin kehitys
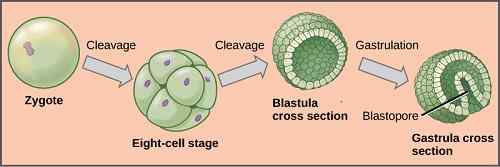
Kun tsygootti on muodostunut, tämä alkaa kokea sarjan muutoksia ja muutoksia, jotka koostuvat peräkkäisestä mitoosisarjasta, jotka muuttavat sen massaksi diploidisoluiksi, jotka tunnetaan nimellä Morula.
Zygootin ylittävä kehitysprosessi kattaa useita vaiheita: segmentointi, räjähdys, maha ja organogeneesi. Jokaisella heistä on enempää merkitystä, koska heillä on avainasemassa uuden olennon muodostumisessa.
-Segmentointi
Tämä on prosessi, jolla zygootti kokee paljon mitoottisia jakautumisia kertomalla sen solujen lukumäärän. Jokainen näistä jakoista muodostuneista soluista kutsutaan blastomeeriksi.
Prosessi tapahtuu seuraavasti: tsygootti on jaettu kahteen soluun, puolestaan nämä kaksi jaetaan aiheuttamalla neljä, nämä neljä kahdeksassa, olet 16 -vuotias ja lopuksi olet 32 -vuotias.
Kompakti solumassa, joka muodostuu, tunnetaan nimellä Morula. Tämä nimi johtuu siitä, että sen ulkonäkö on samanlainen kuin BlackBerry.
Nyt, vitalluksen määrästä ja sijainnista riippuen, segmentointityyppiä on neljä tyyppiä: holoblastinen (kokonaismäärä), jotka voivat olla yhtä suuret tai epätasa -arvoiset; ja meroblastinen (osittainen), joka voi myös olla yhtä suuri tai epätasainen.
Holoblastinen tai kokonaissegmentointi
Tämän tyyppisessä segmentoinnissa koko zygootti on segmentoitu mitoosin kautta, mikä johtaa blastomeereihin. Nyt holoblastinen segmentointi voi olla kahden tyyppistä:
- Sama holoblastinen segmentointi: Tämän tyyppisessä holoblastisessa segmentoinnissa kaksi ensimmäistä jakoa ovat pitkittäisiä, kun taas kolmas on päiväntasaaja. Tämän vuoksi muodostuu 8 blastomeeriä, jotka ovat yhtä suuret. Nämä puolestaan jaetaan mitoosin kautta morulan muodostamiseksi. Holoblastinen segmentointi on tyypillistä isolekyylimunoille.
- Epätasainen holoblastinen segmentointi: Kuten kaikessa segmentoinnissa, kaksi ensimmäistä divisioonaa ovat pitkittäisiä, mutta kolmas on leveys. Tämäntyyppinen segmentointi on tyypillistä mesolekyylimunoille. Tässä mielessä blastomeerit muodostuvat koko tsygootissa, mutta ne eivät ole samoja. Sygootissa, jossa Vitleä on vähän, muodostetut blastomeerit ovat pieniä ja tunnetaan mikrometreinä. Päinvastoin, runsaasti Vitleä sisältävän zygootin osassa syntyvät blastomeerit kutsutaan makromeereiksi.
Meroblastinen tai osittainen segmentointi
Se on tyypillistä tsygootteille, jotka sisältävät runsaasti Vitleä. Tämän tyyppisessä segmentoinnissa vain niin kutsuttu eläinnapa on jaettu. Vegetatiivinen napa ei ole mukana divisioonassa siten, että suuri määrä villusia ei ole segmentointia. Samoin tämäntyyppinen segmentointi luokitellaan diskoidiseksi ja pinnalliseksi.
Meroblastinen diskidinen segmentointi
Tässä kokee vain zygootin eläinnavan. Loput tästä, joka sisältää melko Vitleä, ei ole segmentoitu. Samoin muodostuu blastomeerilevy, joka myöhemmin aiheuttaa alkion. Tämäntyyppinen segmentointi on tyypillistä telolekyysien tsygootteille, etenkin lintuissa ja kaloilla.
Pinnallinen meroblastinen segmentointi
Pinnallisessa meroblastisessa segmentoinnissa ytimessä on useita jakautumisia, mutta sytoplasma ei. Tällä tavoin saadaan useita ytimiä, jotka liikkuvat kohti pintaa, jakautuen koko sytoplasman kannen koko. Myöhemmin solurajoitukset, jotka tuottavat perifeerisesti ja ympäröivää blastodermia, joka ei ole segmentoitu, ilmestyy. Tämäntyyppinen segmentointi on tyypillistä niveljalkaisille.
-Räjähdys
Segmentointia seuraa prosessi. Tämän prosessin aikana blastomeerit liittyvät toisiinsa muodostaen hyvin läheiset ja kompaktit soluyhdistykset. Blastoinnin kautta muodostuu blastula. Tämä on ontto rakenne pallon muodossa, sisäisen ontelon kanssa, joka tunnetaan nimellä Blastocle.
Blastun rakenne
Blastoderma
Se on ulkoinen solukerros, jota kutsutaan myös trofoblastiksi. Se on elintärkeää, koska sen perusteella muodostuu istukka ja napanuora, tärkeät rakenteet, joiden kautta äidin ja sikiön välinen vaihto vahvistetaan.
Se voi palvella sinua: Colima Flora ja eläimistöSen muodostuu suuri joukko soluja, jotka siirtyivät Morulan sisäpuolelta reuna -alueelle.
Blastokle
Se on blastokystin sisäontelo. Se muodostuu, kun blastomeerit siirtyvät morulan ulkoisiin osiin muodostaakseen blastodermin. Räjähdyksen miehittää nesteen.
Embrioblasto
Se on sisäisen solun massa, joka sijaitsee blastosystin sisällä, erityisesti yhdessä sen päistä. Alkiosta alkio itse muodostetaan. Alkion puolestaan koostuu:
- Hypoblast: Solukerros, joka sijaitsee primaarisen Vittelino -säkin ääreisosassa.
- Epiblast: solukerros, joka on amnioottisen ontelon vieressä.
Sekä epiblastiset että hypoblast.
Gastointi
Tämä on yksi tärkeimmistä prosesseista, jotka tapahtuvat alkion kehityksen aikana, koska se sallii kolmen itävän kerroksen muodostumisen: endoderma, mesodermi ja ektoderma.
Se, mitä tapahtuu mahalaukun aikana. Siten, että he siirtyvät kohti hypoblastia, jopa siirtämällä joitain tämän soluja. Siten muodostuu SO -nimeämä primitiivinen viiva.
Välittömästi tapahtuu invaginaatio, jonka läpi kyseisen primitiivisen viivan solut johdetaan Blastoclen suuntaan. Tällä tavoin muodostuu ontelo, joka tunnetaan nimellä Archeistron, jolla on aukko, blastoporo.
Näin muodostuu bilaminar -alkio, joka koostuu kahdesta kerroksesta: endoderma ja ektoderma. Kaikki elävät olennot eivät kuitenkaan tule bilaminar -alkiosta, mutta on muitakin, kuten ihmistä, jotka tulevat trilaminar -alkiosta.
Tämä trilaminaarinen alkio muodostuu, koska archentronisolut alkavat lisääntyä ja jopa sijoittaa ektoderman ja endodermin välillä, mikä aiheuttaa kolmannen kerroksen, mesodermin.
Endodermi
Tästä itävästä kerroksesta muodostuu hengitys- ja ruuansulatusjärjestelmien elinten epiteeli, samoin kuin muut elimet, kuten haima ja maksa.
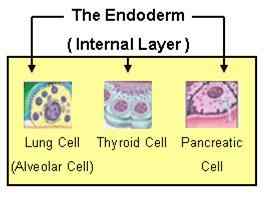 Elimiä, jotka ovat peräisin endodermista. Lähde: endoderm2.PNG: J.Steinbockmaga [julkinen alue]
Elimiä, jotka ovat peräisin endodermista. Lähde: endoderm2.PNG: J.Steinbockmaga [julkinen alue] Mesoderma
Se aiheuttaa luita, rustoa ja vapaaehtoisia tai nauhoitettuja lihaksia. Samoin muodostuu siitä verenkiertoelimet, ja muut, kuten munuaiset, sukurauhaset ja sydänlihakset, muun muassa, muun muassa.
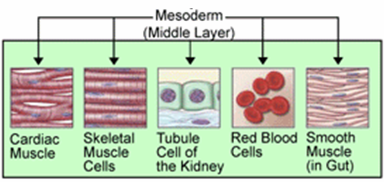 Mesodermista johdetut kankaat. Lähde: J.Steinbock [julkinen alue]
Mesodermista johdetut kankaat. Lähde: J.Steinbock [julkinen alue] Ektoderma
Hän on vastuussa hermoston, ihon, kynsien, rauhasten (hiki ja talirauhasen) muodostumisesta, lisämunuaisesta ja aivolisäkeestä.
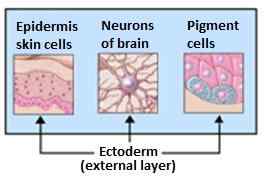 Ektodermin johdannaiset. Lähde: Ecodderm.PNG: Catmaga [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]]
Ektodermin johdannaiset. Lähde: Ecodderm.PNG: Catmaga [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]] Organogeneesi
Se on prosessi, jolla itää ja muutossarjan kautta jokainen elimestä saadaan jokainen elin.
Yleisesti ottaen, mitä täällä organogeneesissä tapahtuu, on se, että kantasolut, jotka ovat osa itäviä kerroksia.
Elävän olennon evoluutiotasosta riippuen organogeneesiprosessi on enemmän tai vähemmän monimutkainen.
Viitteet
- Carrillo, D., Yaser, l. ja Rodríguez, n. (2014). Alkion kehityksen peruskäsitteet lehmässä. VACA: n lisääntyminen: Didaktinen käsikirja lisääntymisessä, raskaudessa, imetyksessä ja naudan naisen hyvinvoinnissa. Antioquian yliopisto. 69-96.
- Cruz, r. (1980). Ihmisen elämän alun geneettinen perusta. Chileläinen lastenlääketieteellinen aikakauslehti. 51 (2). 121-124
- López, c., Garcia, v., Mijares, j., Domínguez, J., Sánchez, f., Álvarez, I. Ja Garcia, V. (2013). Gastointi: avainprosessi uuden organismin muodostumisessa. Asebir. 18 (1). 29-41
- López, n. (2010). Lajimme zygootti on ihmiskeho. Henkilö ja bioetiikka. 14 (2). 120-140.
- Sadler, t. (2001). Langmanin lääketieteellinen embryologia. Pan -american lääketieteellinen toimitus. 8. painos.
- Ventura, P. Ja Santos, M. (2011). Uuden ihmisen elämän alku tieteellisestä biologisesta näkökulmasta ja sen bioeettiset vaikutukset. Biologinen tutkimus. 44 (2). 201-207.
- « Kermésin alkuperä, pelit ja perinteet Latinalaisessa Amerikassa
- Myoosin syyt, patofysiologia ja hoidot »