

Citokroma C -oksidaasirakenne, toiminnot, estäjät
- 1189
- 247
- Eddie Hackett
Se Citokroma C -oksidaasi Se on entsymaattinen proteiinikompleksi, joka voi ylittää solukalvon lipidikerroksen. Se on, että olemme liitetty pääasiassa mitokondrioiden sisäiseen kalvoon, koska ne ovat sekä prokaryoottisissa organismeissa (bakteereissa), kuten eukaryooteissa (yksisoluinen ja monisoluinen).
Tämä entsyymi kutsutaan myös kompleksiksi IV, ja se on elintärkeä organismien aerobisissa metabolisissa toiminnoissa, koska se on välttämätöntä elektronien kuljetusketjussa, jossa solu polttaa sokereita ja vangitsee osan energiasta, joka vapautuu adenosiinin tryfosfaatin tai ATP: n varastointiin.
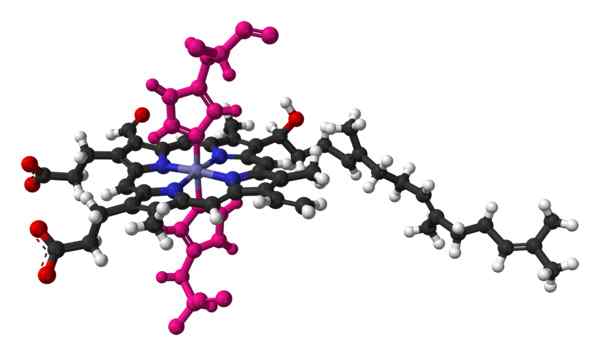
Hemon bola ja palo -malli, molekyyli, joka löytyy naudan sydämen sytokromi c -oksidaasin kiteisestä rakenteesta. Otettu ja muokattu osoitteesta: Benjah-BMM27 [julkinen alue]. Sytokromin nimi tarkoittaa "solpigmenttejä". Nämä ovat veriproteiineja, jotka kuljettavat elektroneja. Irlantilainen lääkäri Charles Alexander MacMunn löysi sytokromit vuonna 1884. Macmunn oli edelläkävijä havaitaan veren hengityspigmentti, nimeltään tänään sytokromi 1.
Viime vuosisadan 20 -luvulla venäläinen entomologi ja parasitologi. Vaikka MacMunn oli löytänyt ne vuonna 1884, tiedeyhteisö oli unohtanut ja jopa jotkut olivat ymmärtäneet heidän työnsä väärin.
[TOC]
Yleiset luonteenpiirteet
Yleensä hengityspigmenteillä on ominaisia näkyviä spektrejä. On tiedossa, että on ainakin neljä kattavaa membraaniproteiinikompleksia, joissa sytokromeja on 5 erityyppistä: A, A3, B, C1 ja C, luokiteltu spektrin absorptio -maksimien aallonpituuksien mukaan.
Niitä löytyy yleensä mitokondrioiden sisäisestä kalvosta. Niitä on kuitenkin havaittu myös endoplasmisessa retikulumissa ja kloroplastit, eukaryoottisissa ja prokaryoottisissa organismeissa.
He esittävät Hemo -proteesiryhmän, joka sisältää rautaa (usko). Jokainen tunnetuista sytokromeista toimii multientsimaattisissa komplekseissa hengitysketjun elektronien kuljetuksessa.
Sytokromeilla on toiminta osallistua oksidin vähentämisreaktioihin. Pelkistysreaktiot, joissa ne hyväksyvät elektroneja, esiintyvät eri tavalla jokaiselle sytokromityypille, ja niiden arvo määritetään hengitysketjun elektronien virtauksella.
-Sytokromi c
Neljä tyyppiä sytokromi c tunnetaan, jotka ovat seuraavat.
Luokka I
Tässä luokassa ovat sytokromit c matalan käännöksen (alamäki), jotka ovat läsnä bakteereissa ja mitokondrioissa. He ovat oktaedrit. Hemo Unionin paikka on histidiinin lopussa ja kuudes ligandi toimitetaan metioniinitähteellä lopussa C.
Tästä luokasta voit jopa tunnistaa useita alaluokkia, jotka on määritetty niiden kolmen dimensioisen rakenteen.
Luokka II
Tässä luokassa on sytokromeja C korkean käännöksen (korkea. Korkea käännös on sitoutumiskohta lähellä pääte-C: tä, ja kuudes ligandi näyttää olevan matalalla käännöksellä olevan metioniinitähde lähellä N-Termus-päätä englanniksi). Ne on pentakoordinoitu viidennellä ligandin histidiinillä.
Voi palvella sinua: turgiteetti (biologia)Luokka III
Tälle luokalle on ominaista sytokromit C, jossa on useita hemo (C3 ja C7) ja matala potentiaalinen oksidi-reduktori, vain 30 aminohappotähteellä hemoa kohden. Tämän luokan edustajilla HEMO C -ryhmillä on rakenne- ja ei -ekvivalenttitoiminnot erilaisten redox -potentiaalien lisäksi. He ovat oktaedrit.
Luokka IV
Joidenkin kirjoittajien mukaan tämä luokka luotiin vain muun muassa muiden proteesiryhmien, samoin kuin Hemo C: n tai flavocytokrome C.
Sytokromi c -oksidaasi tai kompleksi IV
Sytokromi c -oksidaasi on mitokondriaalinen entsyymi, joka suorittaa elektronisen kuljetuksen viimeisen vaiheen solujen hengityksessä. Tämä entsyymi katalysoi elektronien kuljetusta vähentyneestä sytokromi c: stä happea.
Jotkut kemialliset yhdisteet, kuten syanidi, hiilidioksidi ja atsidi. Muut IV -kompleksin estämisen muodot ovat geneettisiä mutaatioita.
Evoluutio -kannalta sytokromi c -oksidaasia löytyy vain aerobisista organismeista, ja useat tutkijaryhmät viittaavat siihen, että tämän proteiinin läsnäolo osoittaa evoluutiosuhteita, joissa kasvit, sienet ja eläimet jakautuvat myös esi -isänsä yhteisesti.
Rakenne
Sytokromi c -oksidaasi muodostaa homodimeerisen kompleksin, toisin sanoen kahdesta samanlaisesta monomeeristä mitokondrioiden sisäisessä kalvossa. Entsymaattinen kompleksi koostuu 3 - 4 alayksiköstä prokaryoottisissa organismeissa ja enintään 13 (jotkut ehdottavat 14) polypeptidiä organismeissa, kuten nisäkkäissä.
Näissä organismeissa 3 polypeptidit ovat mitokondrioiden alkuperää ja loput ovat peräisin ytimestä. Jokaisessa monomeerissä on 28 kalvon läpäisevää potkuria, jotka erottavat hydrofiiliset domeenit membraanimatriisista ja membraanin välisestä tilasta.
Siinä on vain yksi katalyyttinen yksikkö, jota löytyy kaikista entsyymeistä, jotka katalysoivat hapettumista/pelkistysreaktioita, käyttämällä molekyylin happea (oksidaasit, erityisesti Hemo-Cobre). Kompleksi sisältää sytokromit A ja A3 yhdistävät alayksikkö I ja kaksi kuparikeskusta.
Se esittelee yhden tai useamman HEMO C -ryhmän, joka on kytketty ympäröivään proteiinirakenteeseen yhdellä tai useammalla (yleensä kahdella) Tioreter -linkillä. Muut kirjoittajat ehdottavat, että porfyriinirenkaan ja kahden kysteiinitähteen väliseen proteiiniin on kytketty yksi Hemo C -ryhmä.
Ainoa edellä mainitun hemo C -ryhmän ympäröimä on hyodrofobinen jäte ja se on heksakoordininoitu, koska se on histidiini polypeptidiketjun ja menetelmän asemassa 18 80.
Voi palvella sinua: kasvisolu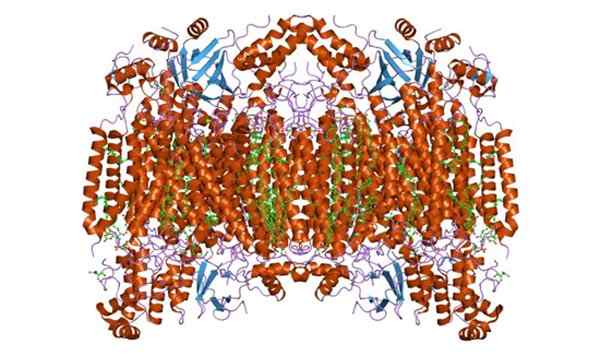 Sytokromi c oksidaasin alayksikkö F. Otettu ja toimitettu: Jawahar Swaminathan ja MSD: n henkilökunta Euroopan bioinformatiikan instituutissa [julkinen verkkotunnus]
Sytokromi c oksidaasin alayksikkö F. Otettu ja toimitettu: Jawahar Swaminathan ja MSD: n henkilökunta Euroopan bioinformatiikan instituutissa [julkinen verkkotunnus] Funktiot
Citokroma C -oksidaasit ovat päähenkilöitä kolmessa primaarisessa fysiologisessa mekanismissa, jotka näemme alla.
Apoptoosi tai ohjelmoitu solukuolema
Apoptoosi on saman organismin aiheuttama. Tässä fysiologisessa prosessissa sytokromi c -oksidaasi osallistuu välittäjäksi.
Tämä mitokondrioiden vapauttama proteiini johtaa vuorovaikutukseen endoplasmisen retikulumin kanssa, mikä aiheuttaa kalsiumin erityksen tai vapautumisen. Kalsiumin asteittainen kasvu laukaisee sytokromi c -oksidaasin massiivisen vapautumisen, kunnes on mahdollista saavuttaa sytotoksiset kalsiumpitoisuudet.
Sytotoksiset kalsiumpitoisuudet ja sytokromien C: n vapautuminen aiheuttavat useiden entsyymien kaskadin aktivaatiota, jotka ovat vastuussa solujen tuhoamisesta.
Solun tai kudoksen uudistaminen
Useat tutkimukset osoittavat, että kun sytokromi c -oksidaasi altistetaan 670 nanometrin aallonpituuksille, osallistuu funktionaaliseen kompleksiin, joka tunkeutuu vaurioituneisiin tai loukkaantuneisiin kudoksiin ja lisää solujen uudistamisen nopeutta.
Energian aineenvaihdunta
Tämä on ehkä sytokromi c -oksidaasin tunnetuin ja merkityksellisin tehtävä. Juuri juuri oksidaasikompleksi (hengitysketjun) oksidaasikompleksi, joka vastaa sytokromi c: n elektronien keräämisestä ja siirtää ne happimolekyyliin, vähentäen sen kahteen vesimolekyyliin,.
Tähän prosessiin kytkettynä protoneihin on siirretty membraanin läpi, mikä johtaa sähkökemiallisen gradientin muodostumiseen, jota ATP -synteesikompleksi käyttää ATP: n tuottamiseen tai syntetisointiin (adenosín triffisphyte).
Estäjät
Citokroma C -oksidaasia estävät useat kemialliset prosessit ja yhdisteet. Tapa, jolla se tapahtuu.
Azidin läsnä ollessa. Tämä aiheuttaa keskeytyksen solujen hengitysprosessissa ja aiheuttaa siksi solujen kemiallisen asfikaatiota.
Muut yhdisteet, kuten typpioksidi, rikkivety, metanoli ja jotkut metyloidut alkoholit, aiheuttavat myös sytokromi c -oksidaasin estämistä.
Puute
Sytokromi c -oksidaasi on entsyymi, jota säätelevät sekä ytimen että mitokondrian geenit. On geneettisiä muutoksia tai mutaatioita, joilla voi olla sytokromi c -oksidaasin puute.
Nämä mutaatiot häiritsevät entsyymin toiminnallisuutta, koska ne muuttavat entsymaattista rakennettaan tuottaen mukanaan aineenvaihduntahäiriöitä alkion kehityksen aikana (ihmisten tutkimusten mukaan), mikä vaikuttaa myöhemmin organismiin ensimmäisinä elämänvuosina.
Voi palvella sinua: Dendriittisolut: Ominaisuudet, toiminto, tyypitSytokromi C -oksidaasin puute vaikuttaa kudoksiin, joilla on suuri energian kysyntä, kuten sydän, maksa, aivot ja lihakset. Näiden mutaatioiden oireet heijastuvat ennen kahta elämää ja voivat ilmetä vahvoina tai lievinä olosuhteissa.
Lieviä oireita voidaan nähdä jopa pian ensimmäisen vuoden jälkeen, ja yleensä heillä on vain lihaksen jännitys (hypotonia) ja lihasten surkastuminen (myopatia) vähenee vain lihasjännitystä (hypotonia).
Toisaalta henkilöillä, joilla on voimakkaampia oireita, voi olla lihasten surkastuminen ja enkefalyopatia. Muut sytokromi c -oksidaasin puuttumisen aiheuttamat olosuhteet ovat hypertrofinen sydänlihaksen, patologinen kasvu maksan koon, Leight -oireyhtymän ja maitohappoasidoosin lisääntyminen.
Käytetään fylogeny
Fylogeny on tiede, joka vastaa alkuperä-, koulutuksen ja evoluutiokehityksen tutkimuksista esi-isien jälkeläisen näkökulmasta, organismeista. Viime vuosikymmeninä fylogene -tutkimukset molekyylianalyysillä ovat olleet yhä yleisempiä, heittäen paljon tietoa ja ratkaisemalla taksonomisia ongelmia.
Tässä mielessä jotkut fylogeneettiset tutkimukset osoittavat, että sytokromi c -oksidaasien käyttö voi auttaa luomaan evoluutiosuhteita. Tämä johtuu siitä, että tämä proteiinikompleksi on erittäin säilynyt ja sitä esiintyy monenlaisissa organismeissa, yksisoluisista protisteista, suuriin selkärankaisiin.
Esimerkki tästä, ovat ihmisillä tehdyt testit, simpanssit (Leipäleipä) ja reesusmakakit (Mulatta Macaca-A. Tällaiset testit paljastivat, että ihmisten sytokromi c -oksidaasimolekyylit ja simpanssit olivat identtisiä.
Hän osoitti myös, että reesuksen makakin sytokromi c -oksidaasimolekyylit.
Viitteet
- R -.P. Ambler (1991). Bakteerikykroomien sekvenssivaihtelu c. Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- Sytokromi c. Toipunut NewworldyClopediasta.org.
- V.Colman, E. Kustannus. Chaves, v. Tortora (2015). Sytokromi C: n biologiset roolit: mitokondriaalinen elektroninen kuljetus, ohjelmoitu solukuolema ja AUS -aktiivisuusvoitto. Lääketieteellisen tiedekunnan vuosipäivä.
- Sytokromi c oksidaasin alayksikkö I. Toipunut EBI: stä.Ac.Yhdistynyt kuningaskunta.
- Lens. Youfen, P. Jeong-Oon, D. Jian-Hong & B. Yidong (2007). Sytokromi C -oksidaasin alayksikkö IV on välttämätöntä entsyymikompleksin kokoonpano- ja hengityselimille. Journal of Bioenergetics and Biomembranes.
- Geeniryhmä: Kompleksi IV mitokondriaalinen: sytokromi c oksidaasi-alayksiköt (Cox, MT-CO). Toipunut genenimistä.org.
- JA.F. Hartree (1973). Sytokromin löytäminen. Biokemiallinen koulutus.
- Citokroma C -oksidaasi, .. . Salatuna Ivami.com.
- C.K -k -. Mathews, k.JA. Pakettiauto Hold & k.G. Ahern (2002). Biokemia. 3. painos. Benjamin / Cummings Publishing Company, Inc.
- « Opotillo -ominaisuudet, elinympäristö, ominaisuudet ja tuholaiset
- Pinus patula -ominaisuudet, elinympäristö, taksonomia, käyttö, tuholaiset »