

Sytotrofoblastiominaisuudet, kehitys ja toiminta
- 4755
- 1491
- Edgar VonRueden
Hän sytotrofoblasti o Langhans -solut, se on osa mononukleisoituneista soluista koostuvan trofoblastien peruskalvosta. Tämä osa vastaa kantasolujen populaatiota, josta muut trofoblastit johtuvat.
Tämä solukerros mitoottisesta näkökulmasta on erittäin aktiivinen, tuottaen soluja, jotka sitoutuvat synkytiotrofoblastiin. Sytotrofoblasti on peräisin blastosystin implantoinnin ajanjaksosta nisäkkäiden alkion kehityksessä. Tämän kehitysvaiheen aikana trofoblastiset solut lisääntyvät antaen tietä hyökkäykseen endometriumin epiteelissä.
Lähde: Henry Vandyke Carter [julkinen alue] [TOC]
Ominaisuudet
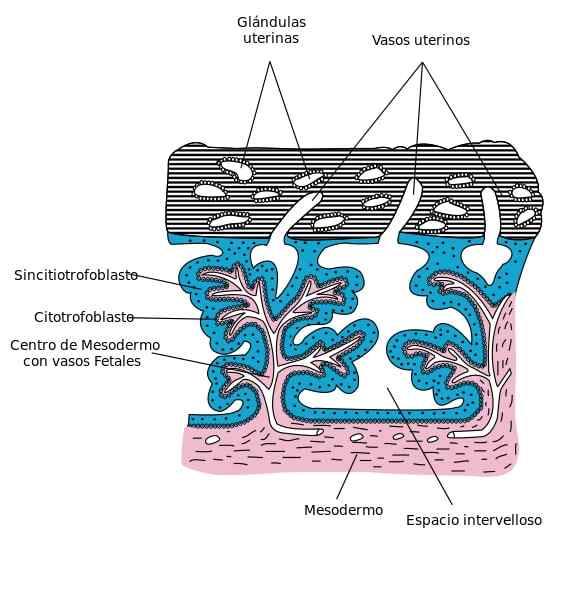
Kerros mononukleoituja soluja muodostavat sytotrofoblastien, trofoblastien sisäpuolelle. Nämä solut sijaitsevat koorion villissä, ja ne peittävät synkronoinnin. Sytotrofoblastille on tunnusomaista korkea solujen erilaistuminen ja lisääntymiskyky sekä alhainen funktionaalinen aktiivisuus.
Alkion implantaation vaiheen tai ikkunan aikana sytotrofoblastisolut ovat peräisin kuutiometreistä ja vaaleaksi hyvällä ydinkuvalla ja hyvin erilaistuneet toisistaan.
Sytotrofoblastisten solujen lisääntyminen tapahtuu jatkuvalla solujen kertolaskulla. On arvioitu, että ainakin puolet tämän kerroksen soluista suorittaa solusyklin. Proliferaation lisäksi on korkea soluerottelu, joka tuottaa synkytiotrofoblastien makaa.
Extravellliterian sytofoblast
Sytofoblastien ekstraveloiva. Tämä solukerros lisääntyy nopeasti tunkeutuvan kohdun strooman ja endometriumin kierrevaltimoiden vähentämällä verisuonien seinien vastustuskykyä. Kaksi tyyppiä ekstravellit -sytofoblastityyppejä erotetaan: interstitiaalit ja endovaskulaariset.
Interstitiaalissa solut tunkeutuvat myometriumiin sulautumaan ja muuttumaan suuriksi istukan soluiksi. Nämä solut eivät tunkeudu verisuoniseiniin.
Voi palvella sinua: PachyneToisaalta endovaskulaari tunkeutuu verisuoniseinämiin, jotka tuhoavat verisuonen keskikerroksen sileät solut, ja siinä on sen sijaan fibrinoidimateriaali. Endoteelifenotyyppiä simuloivien molekyylien diffuusio mahdollistaa äidin verisuonten endoteelin korvaamisen uudella sisäpinnalla.
Sytotrofoblasti -aktiivisuutta säätelevät geneettiset, transkriptiot, kasvu, hormonaaliset ja kemialliset tekijät (kuten molekyylin happipitoisuus).
Kehitys ja toiminta
Nisäkkäissä, kun munasolun hedelmöitetään siittiöitä, suoritetaan sarja solujakoja, kunnes muodostuu blastosysti, joka on onto Alkion kudokset kutsuvat alkion.
Blastosysti on kiinnitetty endometriumiin toteutuksen aikana. Trofoblastiset solut alkavat levitä, kun ne ovat kosketuksissa endometriumin kanssa, mikä erottaa sytotrofoblastit ja synkytiotrofoblastit.
Ihmislajeissa toteutus tapahtuu suunnilleen kuudenteen päivään munasolun ovulaation ja hedelmöityksen jälkeen.
Joissakin nisäkkäissä tätä vaihetta lykätään päivien, viikkojen tai jopa kuukausien ajan uuden rodun saapumisen välttämiseksi ei -rappeutumishetkellä, kuten ajanjaksoina, jolloin resurssit vähenevät tai kun äiti imettää toista jalostusta.
Eläimissä, kuten karhuissa, testamentissa, sinetöissä ja kamelissa, alkion diapase -nimisen implantaatioikkunan viivästyminen.
Blastosysti pysyy siinä tilassa ilman sytotrofoblastisten solujen lisääntymistä hormonaalisella vaikutuksella. Tämä mekanismi käynnistyy vastauksena ympäristötekijöihin tai pitkittyneelle imetykselle äidissä.
Voi palvella sinua: Mitkä ovat kromoplast?Istukan muodostuminen
Istukka on vastuussa sikiön pitämisestä kehityksessä ja johtuu koorionista (sikiön osuus) ja peruslomakkeesta. Siinä tapahtuu kaasu- ja metaboliittien vaihtoja äidin ja sikiön kierteiden välillä. Tämä elin kehittyy, kun se on peräisin erilaisesta villistä.
Kun sytotrofoblastisolut ja koorionin mesenterian ja verisuonten kehittymisen myötä muodostetaan laajentuvia, primaarisia, toissijaisia ja tertiäärisiä koorionisia villit.
Sytotrofoblasti lisääntyy nopeasti, siirtämällä solut veren laguuniin synkronototrofoblastien sisällä, muodostaen primaarisen koorionin villin.
Myöhemmin nämä villit tunkeutuvat koorionin alkion mesenkyymiin, joka pysyy sytotrofoblastien sisällä ja ympäröimänä, muodostaakseen toissijaisen villin, joka kattaa koorionipussiin.
Tertiäärinen villi muodostuu verisuonten esiintymisen myötä sekundaarisen villin mesenkyymissä.
Syncitiotroplastiin kautta muodostetaan sytotrofoblastien tertiääriset villit, langat tai cupcated -solut.
Tällä tavoin erilaiset solujen agglomeraatit menevät ulkomaille ja liityvät yhteen, peittäen synkronointiantrofoblastien sytotrofoblastisella peitolla. Tämä kansi keskeytetään, missä äidin verisuonet menevät Villositaritiloihin.
Äiti-sikiön rajapinta
Äiti-sikiön rajapinnan ensimmäinen vaihe koostuu ekstravelliterialaisen sytofoblastien (joka sijaitsee istukan villien ulkopuolella) hyökkäyksestä kohdun kierrevaltimoissa, jotka antavat näille valtimoihin korkean tason ominaispiirteet ja niillä on alhainen virtauskestävyys. Tällä tavoin ylläpidetään sikiön kasvun riittävä perfuusio.
Toisessa vaiheessa sytotrofoblastisolut yhdistävät, purkaen solumembraanit, synkrotiotrofoblastien monisydämeen kerroksen aiheuttamiseksi. Jälkimmäinen kääri istukan eriytetyt villit.
Voi palvella sinua: Hepar Will sulfaatti: toiminnot, synteesi, suhde sairauksiinNäiden rajapinnan kahden vaiheen oikea kehitys varmistaa oikea istukka ja siten onnistunut sikiön kehitys ja painovoimatilan turvallinen edistyminen.
Istukan este erottaa äidin ja sikiön veren
Istukan este, joka on pääosin muodostettu sikiön kudoksen kerrokset, vastaa veren erottamisesta äidin veren sikiöstä. Ihmisillä, neljännestä kehityskuukaudesta lähtien, tämä este muuttuu erittäin ohueksi, mikä helpottaa tuotteiden siirtoa sen läpi.
Kuoren tai sytotrofoblastisen peitteen rappeutuminen on syynä istukan esteen painonpudotukseen, jonka rappeutuneessa tilassa se koostuu synkriloivan sytotrofoblastisesta epäjatkuvasta peitteestä, trofoblastisen arkin, samanaikaisuuden mesenkymosin, endoteliumin ja endoteliumin basaalisen lopun, Kolmannen villuksen istukan kapillaarit sikiö.
Istukan este, joka eroaa veren äidistä ja sikiön verestä, on vastuussa hapenvaihdosta ja hiilidioksidista ja metaboliitteista äidin ja sikiön kierteiden välillä.
Viitteet
- Hernández-valencial, m., Valencia-Ortega, J., Ríos-castillo, b., Cruz-Cruz, P. D -d. R -., & Vélez-Sánchez, D. (2014). Implantaation ja istutuksen elementit: kliiniset ja melekulaariset näkökohdat. Meksikon lisääntymislääketieteellinen aikakauslehti, 6 (2), 102-16.
- Mäki, r. W -., Wyse, g. -Lla., Anderson, m., & Anderson, M. (2004). Fysiologinen eläin (Vol. 2). Sunderland, MA: Sinauer Associates.
- Kardong, k. V. (tuhatyhdeksänsataayhdeksänkymmentäviisi). Selkärankaiset: vertaileva anatomia, funktio, evoluutio. Ed. McGraw Hill.
- Rodríguez, M., Couve, c., Egaña, G., & Chamy, V. (2011). Istukan apoptoosi: Molekyylimekanismit preclampsian syntyessä. Chilen synnytys- ja gynekologialehti, 76 (6), 431-438.
- Ross, m. H., & Pawlina, W. (2007). Histologia. Ed. Pan -American Medical.
- Welsch, u., & Sobotta, J. (2008). Histologia. Ed. Pan -American Medical.
- « Syncitiotrofoplasti -ominaisuudet, toiminta, istukan apoptoosi
- Basofiilien ominaisuudet, morfologia, toiminnot, sairaudet »