

Transkriptiotekijän toimintamekanismi, tyypit, toiminnot
- 2073
- 296
- Ronald Reilly
Eräs Transkriptiotekijä Se on geneettiseen transkriptioon tarvittava säätely "lisävaruste" -proteiini. Transkriptio on geneettisen ekspression ensimmäinen vaihe ja merkitsee DNA: n sisältämien tietojen siirtoa RNA -molekyyliin, joka myöhemmin prosessoidaan geenituotteiden aiheuttamiseksi.
RNA -polymeraasi II on entsyymi, joka on vastuussa useimpien eukaryoottisten geenien transkriptiosta ja tuottaa joidenkin pienten RNA: ien lisäksi Messenger -RNA: t, jotka sitten transloidaan proteiineiksi. Tämä entsyymi vaatii tyyppisiä transkriptiotekijöitä, jotka tunnetaan yleisesti tai perustranskriptiotekijöinä.
Nämä eivät kuitenkaan ole ainoita luonnossa esiintyviä transkriptiotekijöitä, koska on olemassa ”ei-yleisiä” proteiineja, sekä eukaryooteissa että prokaryooteissa ja kaareissa, jotka osallistuvat kudosspesifisen geneettisen transkription säätelyyn (monisoluisissa organismeissa tai geeniaktiivisuuden säätelyssä vasteena erilaisille ärsykkeille.
Nämä transkriptiotekijät ovat erittäin tärkeitä efektoreita, ja niitä voidaan löytää käytännössä kaikissa elävissä organismeissa, koska ne edustavat geneettisen ekspression päälähdettä.
Yksityiskohtaiset tutkimukset eri transkriptiotekijöistä erityyppisissä elävissä organismeissa osoittavat, että niillä on modulaarinen rakenne, jossa erityinen alue vastaa vuorovaikutuksesta DNA: n kanssa, kun taas toiset tuottavat stimuloivia tai estävää vaikutusta.
Transkriptiotekijät osallistuvat sitten geneettisten ekspressiokuvioiden mallintamiseen, joilla ei ole mitään tekemistä DNA -sekvenssin muutosten kanssa, mutta epigeneettisten muutosten kanssa. Näiden muutosten tutkimisesta vastuussa oleva tiede tunnetaan nimellä epigenetiikka.
[TOC]
Toimintamekanismi
Toimintojensa käyttämiseksi transkriptiotekijöiden on kyettävä tunnistamaan ja liittämään erityisesti tietty DNA -sekvenssi positiivisesti tai negatiivisesti vaikuttamaan DNA: n mainitun alueen transkriptioon.
Yleiset transkriptiotekijät, jotka ovat periaatteessa samat eukaryooteissa olevien tyypin II geenien transkriptiossa, kootaan ensin geenin promoottorialueelle, mikä ohjaa polymeraasientsyymin ja kaksinkertaisen potkurin ”avaamisen” sijaintia ja "avaamista".
Prosessi annetaan useita peräkkäisiä vaiheita:
- Yleisen transkriptiotekijän TFIID: n liitto toistuvan Timinan (T) ja adeniinin (A) sekvenssiin geenissä, joka tunnetaan nimellä “laatikko Tata”; Tämä aiheuttaa DNA: n vääristymiä, jotka ovat välttämättömiä muiden proteiinien yhdistämiseksi promoottorialueelle.
- Muiden yleisten tekijöiden takaosan kokoonpano (TFIIB, Tfiih, Tfih, Tfiie, TFIIF jne.) ja RNA -polymeraasi II, muodostaen niin kutsutut Transkription aloituskompleksi.
Voi palvella sinua: kromatideja- Aloituskompleksin vapauttaminen, polymeraasin fosforylaatio TFIIH -tekijällä ja RNA -molekyylin transkription ja synteesin alussa transkriptoidun geenin sekvenssistä.
Transkription aktivointi ja tukahduttaminen
Kuten mainittiin, "ei -yleiset" transkriptiotekijät voivat säätää geenien ilmentymistä positiivisesti tai negatiivisesti.
Aktivointi
Jotkut näistä proteiineista sisältävät DNA -liiton rakenteellisten domeenien lisäksi muita syitä, jotka tunnetaan aktivointidomeeneina, joissa on runsaasti happamahappotähteitä, glutamiinia tai proliinitähteitä.
Nämä aktivaatiodomeenit ovat vuorovaikutuksessa yleisen transkriptiotekijöiden kompleksin elementtien kanssa tai niihin liittyvien molekyylien kanssa, jotka ovat suoraan vuorovaikutuksessa kompleksin kanssa. Tämä vuorovaikutus johtaa transkriptiivisen kompleksin kokoonpanon stimulaatioon tai sen aktiivisuuden lisääntymiseen.
Tukahduttaminen
Useimmat transkriptiotekijät estävät transkriptiota häiritsemällä positiivisesti toimivien transkriptiotekijöiden aktiivisuutta estäen sen stimuloivan vaikutuksen. Ne voivat toimia estämällä positiivisen tekijän liiton DNA: lle tai vaikuttamalla tekijöihin, jotka inaktivoivat kromatiinin rakenteen.
Muut estävät tekijät toimivat suoraan estämällä transkription estämättä minkään aktivoivan transkriptionaalisen tekijän vaikutusta; ja vähentää transkription perustasoa vielä alemmalle tasolle kuin se saavutetaan aktivoivien tekijöiden puuttuessa.
Kuten proteiinien aktivointi, tekijöiden tukahduttaminen toimii suoraan tai epäsuorasti perus- tai yleisten transkriptiotekijöiden kanssa.
Kaverit
Vaikka suurin osa transkriptiotekijöistä luokitellaan niiden DNA -union -domeenien ominaisuuksien tai identiteetin mukaan, on joitain, jotka on myös luokiteltu transkriptiotekijöiksi, jotka eivät ole vuorovaikutuksessa suoraan DNA: n kanssa ja tunnetaan transkriptiotekijöinä "epäsuoriksi".
Suorat transkriptiotekijät
Ne ovat yleisimpiä transkriptiotekijöitä. Heillä on DNA -union -domeeneja ja ne voivat aktivoida tai estää geeniekspression liiton avulla spesifisille DNA -alueille. Ne eroavat toisistaan erityisesti DNA -union -domeeniensa ja oligomerointivaltionsa suhteen.
Tämän tyyppisten tekijöiden tutkitut ja tunnustetut perheet ovat:
Helix-Gutero-Hélice ("Kierre-käänne”, HTH)
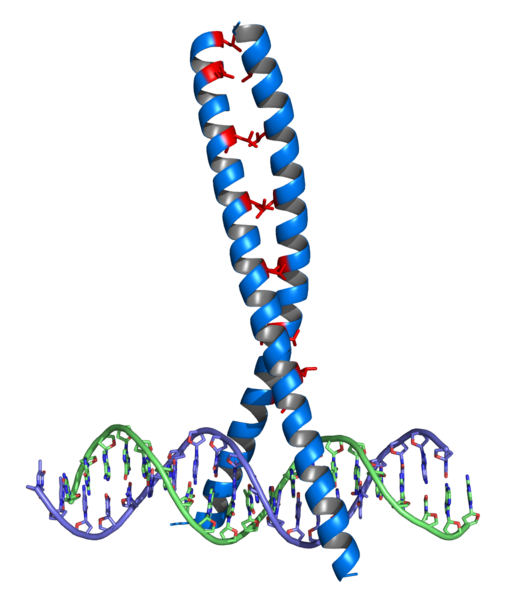
Tämä oli ensimmäinen DNA -union -domeenien tekijöiden perhe, joka löydettiin, ja sitä esiintyy monissa eukaryooteissa ja prokaryooteissa. Sen syyt tunnustamiseen koostuu a -potkurista, käännöksestä ja toisesta potkurista α.
Ne ovat säilyttäneet glysiinin domeenit käännöksen alueella ja myös joitain hydrofobisia jätteitä, jotka auttavat vakauttamaan kahden potkurin järjestelyn HTH -yksikössä.
Voi palvella sinua: geneettinen uima -allasHomeodominium
Sitä esiintyy suuressa määrässä eukaryoottisia säätelyproteiineja. Ensimmäiset sekvenssit tunnistettiin säätelyproteiineissa Drosophila. Tämä domeeni sisältää HTH: n syyn liittyä DNA: hon ja ylimääräiseen a-potkuriin laajennetun N-terminaalisen käsivarren lisäksi.
Sinkkisormit
Löydettiin transkriptiotekijästä TFIIIA Ksenopus Ja on osoitettu, että he osallistuvat moniin eukaryoottisen geneettisen säätelyn näkökohtiin. Niitä löytyy proteiineista, jotka ovat indusoitu erilaistumisen ja kasvun, protoonkogeeneissä ja joissakin yleisissä transkriptiotekijöissä.
Niille on ominaista toistojen läsnäolo sinkin motiiveissa 30 jätettä, joka sisältää useita jätteitä ja histidiinijätettä.
Steroidireseptorit
Tämä perhe sisältää merkittäviä säätelyproteiineja, joilla on domeenin lisäksi hormonien liiton osalta DNA -liiton domeeni ja yleensä ne toimivat transkriptionaalina aktivaattoreina.
Union -domeenit sisältävät 70 jätettä, joista 8 säilytetään kysteiinitähteitä. Jotkut kirjoittajat katsovat, että nämä tekijät voisivat muodostaa sinkkisormien parin, kun otetaan huomioon kaksi neljäskysteinin peliä.
Leucina ja helik-bucle-Herran sulkeminen (""Helix-Loop-Helix "-A
Nämä transkriptiotekijät osallistuvat erilaistumiseen ja kehittämiseen ja heterodimeerin muodostumiseen. Leuciinin sulkemisdomeeni havaitaan erilaisissa eukaryooteissa, ja sille on ominaista kaksi aliverkkotunnusta: dimerisaatiota mittaavien leuciinien sulkeminen ja DNA: n liiton perusalue.
Syyt β Syyt
Niitä esiintyy pääasiassa eukaryoottisista tekijöistä ja eroavat.
Epäsuorat transkriptiotekijät
Tämän tyyppiset transkriptiotekijät vaikuttavat sen sääteleviin vaikutuksiin geneettiseen ilmentymiseen, ei suoran vuorovaikutuksensa kautta DNA: n kanssa, vaan proteiini-proteiini-vuorovaikutuksiin muiden transkriptiotekijöiden kanssa, jotka ovat vuorovaikutuksessa DNA: n kanssa. Siksi heitä kutsutaan "epäsuoriksi".
Ensimmäinen kuvaus oli viruksen trans-aktivaattori "Herpes yksinkertainen " (HSV), joka tunnetaan nimellä VP16, joka sitoutuu OCT-1-tekijään, kun solut ovat tartunnan saaneet tällä viruksella, stimuloimalla spesifisen geenin transkriptiota.
Tämän tyyppiset tekijät, kuten ne, jotka sitoutuvat DNA: han, voivat aktivoida tai tukahduttaa geenitranskription, joten niitä kutsutaan "pakkoviksi" ja "korjattuiksi", vastaavasti.
Säätö
Näitä proteiineja voidaan säädellä kahdella tasolla: niiden synteesissä ja aktiivisuudessaan, mikä riippuu eri muuttujista ja monista tilanteista.
Synteesin säätely
Sen synteesin säätely voi liittyä tiettyjen transkriptiotekijöiden ekspressiokudosspesifisiin. Esimerkki tästä voi olla MYOD -tekijä, syntetisoitu vain luurankoisoluissa ja on välttämätön sen erittelemättömien fibroblastien erilaistumiseen.
Voi palvella sinua: linkitetyt geenitVaikka synteesin säätelyä käytetään pohjimmiltaan geneettisen ekspression hallintaan tietyissä solutyypeissä ja kudoksissa, tämä ei ole ainoa tapa, koska vasteeseen osallistuvien geenien induktioon liittyvien tekijöiden synteesiä säädetään myös useille ärsykkeille.
Toiminnan säätely
Toinen transkriptiotekijöiden säätelymekanismi on sen aktiivisuuden säätely, joka liittyy muiden ennen olemassa olevien transkriptiotekijöiden aktivoitumisen kanssa, joilla on positiivisia tai negatiivisia vaikutuksia tietyn tekijän aktiivisuuteen.
Näiden "sekundaaristen" tekijöiden aktivointi tapahtuu yleensä erilaisilla mekanismeilla, kuten ligandiliitto, proteiini-proteiinin muutokset, fosforylaatiovuorovaikutukset, muun muassa muun muassa.
Toiminnot ja merkitys
Transkriptiotekijät osallistuvat moniin prosesseihin, kuten alkion kehitykseen, kasvuun ja erilaistumiseen, solusyklin hallintaan, sopeutumiseen vaihteleviin ympäristöolosuhteisiin, solujen ja kudosten spesifisten proteiinisynteesimallien ylläpitämiseen jne.
Esimerkiksi kasveissa heillä on tärkeitä toimintoja puolustus- ja vastetapahtumissa erityyppisiin stressiin. On määritetty, että eläinten osteogeneesiä säätelevät transkriptiotekijät, samoin kuin monet muut eri solulinjojen erilaistumisprosessit.
Kun otetaan huomioon näiden proteiinien merkitys organismeissa, ei ole harvinaista ajatella, että näiden sääntelyelementtien muutokset aiheuttavat vakavia patologisia muutoksia.
Ihmisten tapauksessa transkriptiotekijöihin liittyvät patologiat voivat olla kehityshäiriöitä (esimerkiksi transkriptiotekijöiden inaktivoinnin aiheuttamista mutaatioista), hormonaalisen vasteen tai syöpien häiriöt.
Viitteet
- Alberts, b., Dennis, b., Hopkin, k., Johnson, a., Lewis, J., Raff, m.,... Walter, P. (2004). Välttämätön solubiologia. Abingdon: Garland Science, Taylor & Francis Group.
- Iwafuchi-doi, m., & Zaret, K. S. (2014). Pioneer -transkriptiotekijät solujen uudelleenohjelmoinnissa. Geenit ja kehitys, 28, 2679-2692.
- Latchman, D. (1997). Transkriptiotekijät: yleiskatsaus. Int. J -. Biokemia. Solu. Bioli., 29(12), 1305-1312.
- Latchman, D. S. (2007). Transkriptiotekijät. Biotieteiden tietosanakirja, 1-5.
- Marie, P. J -. (2008). Osteoblastogeneesiä säätelevät transkriptiotekijät. Biokemian ja biofysiikan arkistot, 473, 98-105.
- Pabo, c., & Sauer, R. T. (1992). Transkriptiotekijät: rakenneperheet ja DNA: n tunnistamisen periaatteet. Annu. Rev., 61, 1053-1095.
- Singh, k. B -., Foley, r. C., & Oñate-Sánchez, L. (2002). Transkriptiotekijät kasvien puolustuksessa ja stressivasteessa. Nykyinen mielipide kasvibiologiassa, 5, 430-436.
- « Linja -autojen ominaisuudet, ruoka, lisääntyminen, elinympäristö
- Klorofyytin ominaisuudet, elinympäristö, lisääntyminen, ruoka »