

Aerobia -glykolyysi mikä on, reaktiot, glykolyyttiset välittäjät
- 3362
- 201
- Kelly Kilback
Se Aerobinen tai aerobinen glykolyysi Se määritellään glukoosiliikaksi, jota ei käsitellä oksidatiivisella fosforylaatiolla kohti "fermentoivien" tuotteiden muodostumista, jopa korkeiden happipitoisuuksien olosuhteissa ja huolimatta energian suorituskyvyn laskusta.
Se annetaan yleensä kudoksissa, joilla on korkea proliferatiiviset nopeudet, joiden glukoosin ja hapen kulutus on korkea. Esimerkki tästä ovat syöpäkasvainsolut, jotkut nisäkkäiden veren loisolut ja jopa joidenkin nisäkkäiden aivojen solut.
Glukoosin katabolismin uutettu energia säilyy ATP: n ja NADH: n muodossa, joita käytetään alavirtaan eri aineenvaihduntareiteillä.
Aerobisen glykolyysin aikana pyruvaatti on suunnattu Krebs -sykliin ja elektronikuljetinketjuun, mutta sitä käsitellään myös Fermentive NAD+: n uudistamisessa ilman ylimääräistä ATP -tuotantoa, joka päättyy laktaatin muodostumiseen.
Aerobinen tai anaerobinen glykolyysi esiintyy pääasiassa sytosolissa, lukuun ottamatta organismeja, kuten tripanosomatideja, jotka ovat erikoistuneet glykosomiksi tunnetuina glykolyyttiset organelit.
Glykolyysi on yksi tunnetuimmista aineenvaihduntareiteistä. Gustav Embden ja Otto Meyerhof formuloivat sen kokonaan 1930 -luvulla, jotka tutkivat reittiä luuston lihassoluissa. Aerobinen glykolyysi tunnetaan kuitenkin Warburg -vaikutukseksi vuodesta 1924.
Aerobiset glykolyysireaktiot
Glukoosin aerobinen katabolismi esiintyy kymmenessä katalysoidussa vaiheessa. Monet kirjoittajat katsovat, että nämä vaiheet on jaettu energiainvestointivaiheeseen, jonka tavoitteena on lisätä välittäjien vapaata energiapitoisuutta, ja toinen korvaava ja ATP -muotoinen energian voitto.
Voi palvella sinua: Histokemia: perusta, käsittely, värjäysEnergiainvestointivaihe
Glukoosin 1-fosforylaatio glukoosiksi 6-fosfaatiksi, jota katalysoidaan heksakinaasilla (HK). Tässä reaktiossa se sijoitetaan jokaiselle glukoosimolekyylille, ATP -molekyylille, joka toimii fosfaattiryhmän luovuttajana. Glukoosi 6-fosfaatti (G6P) ja ADP, ja reaktio on peruuttamaton.
Entsyymi vaatii täydellisen Mg-ATP2-.
G6P: n 2-assomerointi fruktoosiksi 6-fosfaatiksi (F6P). Siinä ei ole energiankulutusta ja se on palautuva reaktio, jota katalysoi fosfoglukoosi -isomeraasi (PGI).
3-F6P-fosforylaatio fruktoosiin 1,6 biffosfaattia katalysoi fosfofratokinaasi-1 (PFK-1) (PFK-1). ATP-molekyyliä käytetään fosfaattiryhmän luovuttajana ja reaktiotuotteet ovat F1.6-BP ja ADP. ∆G: n arvon ansiosta tämä reaktio on peruuttamaton (reaktio 1).
F11.6-bp: n 4-katalyyttiset kopiot dihydroksiasetonin fosfaatissa (DHAP), ketoosi ja glyseraldehydi 3-fosfaatti (GAP), aldosa. Aldolaasientsyymi on vastuussa tästä palautuvasta aldoolisesta kondensaatiosta.
5-Triosa-fosfaatti-isomeraasi (TIM) on vastuussa triosasfosfaatin: DHAP: n ja aukon kääntämisestä ilman ylimääräistä energiaa koskevaa vaikutusta.
Energian talteenottovaihe
1-the Gap hapettaa glyseraldehydi 3-fosfaattidehydrogenaasi (GAPDH), joka katalysoi rakofosfaattiryhmän siirtoa muodostamaan 1,3-bifoscglycererate. Tässä reaktiossa kaksi NAD+ -molekyyliä vähenee glukoosimolekyylillä, ja käytetään kahta epäorgaanista fosfaattimolekyyliä.
Jokainen tuotettu NADH kulkee elektronikuljetinketjun läpi ja 6 ATP -molekyyliä syntetisoidaan oksidatiivisella fosforylaatiolla.
2-fosfoglysererate-kinaasi (PGK) siirtää fosforyliryhmän 1,3-bifosfoglyseraatiosta ADP: hen, muodostaen kaksi ATP-molekyyliä ja kaksi 3-fosfoglyseraatista (3PG) (3PG). Tätä prosessia kutsutaan fosforylaatioksi substraatin tasolla.
Voi palvella sinua: Stabilisaattorin valinta: Mikä on ja esimerkkejäPGK vastaa kahteen HK- ja PFK -reaktioihin kulutettuihin ATP -molekyyleihin tällä reitin kuluessa PGK.
3--3PG muunnetaan 2pg: ksi Mutasa-fosfoglyktiona (PGM), joka katalysoi fosforyyliryhmän siirtymistä glysereen hiilen 3 ja 2 välillä kahdessa vaiheessa ja palautuvasti. Tämä entsyymi vaati myös magnesium -ionia.
4-A-dehydraatioreaktio, jota katalysoi enolasas, muuntaa 2PG fosfoenolypyruvaatiksi (PEP) reaktioksi, joka ei vaadi energiainvestointeja, mutta tuottaa yhdisteen, jolla on suurempi energiapotentiaali fosfaattiryhmän siirtämiseksi myöhemmin.
5-Finaalisesti kinaasi-pyruvaatin (PYK) katalysssi. Glukoosimolekyyliä käytetään kahta ADP -molekyyliä ja syntyy 2 ATP -molekyyliä. PYK käyttää kalium- ja magnesiumioneja.
Siten glykolyysin kokonaisenergian suorituskyky on 2 ATP -molekyyliä jokaiselle reitille kulkevalle glukoosimolekyylille. Aerobisissa olosuhteissa täydellinen glukoosin hajoaminen merkitsee 30 - 32 ATP -molekyylin saamista.
Glukoliittisten välittäjien kohde
Sitten glykolyysi, pyruvaatti altistetaan dekarboksylaatiolle, tuottaen CO2: ta ja luovuttaa asetyyliryhmä asetyylikoentsyymiin A, joka myös hapettuu CO2: lle Krebs -syklissä.
Tämän hapettumisen aikana vapautuvat elektronit kuljetetaan happea mitokondrioiden hengitysketjun reaktioiden kautta, mikä lopulta edistää ATP: n synteesiä tässä organelissa.
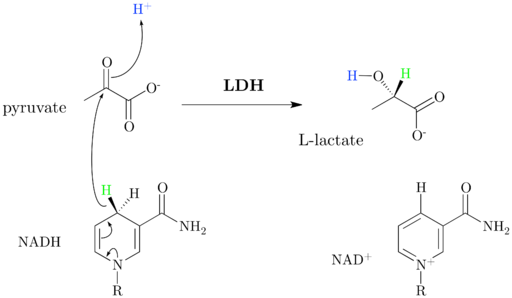
Se voi palvella sinua: Flora ja Efauna de Salta: Edustavampi lajiAerobisen glykolyysin aikana tuotettu ylimääräinen pyruvaatti prosessoidaan laktaattidehydrogenaasin entsyymillä, joka muodostaa laktaattia ja regeneroi osan NAD+: sta kulutettujen askelten glykolyysissä, mutta ilman uusien ATP -molekyylien muodostumista.
Laktaattidehydrogenaasimekanismi (lähde: jazzlw [CC BY-Sa 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta) Lisäksi pyruvaattia voidaan käyttää anabolisissa prosesseissa, jotka johtavat esimerkiksi alaniinin aminohapon muodostumiseen tai voivat toimia myös rasvahappojen synteesin luurankoina.
Kuten pyruvaatti, glykolyysin lopputuote, monet reaktion välittäjistä täyttävät muut toiminnot katabolisilla tai anabolisilla reiteillä, jotka ovat tärkeitä solulle.
Näin on glukoosi 6-fosfaatin ja pentoosifosfaatin reitti, missä saadaan.
Viitteet
- Akram, m. (2013). Mini-katsaus glykolyysistä ja syövästä. J -. Syöpä. Koulutus., 28, 454-457.
- ESEN, E., & Pitkä, f. (2014). Osteoblastien aerobinen glykolyysi. Curr Osteoporos Rep, 12, 433-438.
- « Efebofilia Mikä on, oireet, syyt, seuraukset, hoito
- Isomeraasi mikä on, toiminnot, nimikkeistö, tyypit »