

Megacariocites -ominaisuudet, rakenne, koulutus, kypsä
- 4914
- 606
- Dr. Travis Reichert
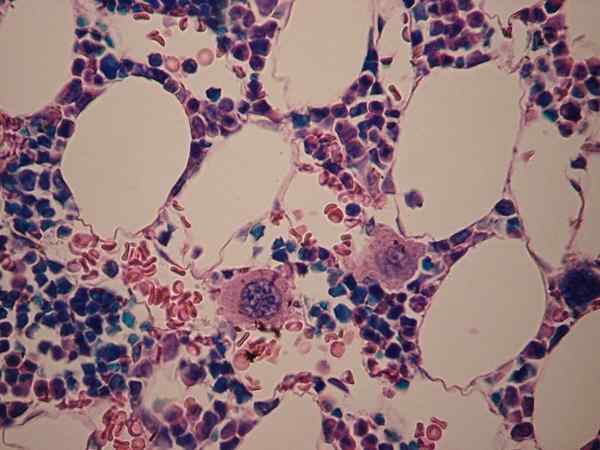
Se Megakariosyyttit Ne ovat huomattavan kokoisia soluja, joiden solujen pirstoutuminen aiheuttaa verihiutaleita. Kirjallisuudessa niitä pidetään "jättiläisinä" soluina, jotka ylittävät 50 um, joten ne ovat hematopoieettisen kudoksen suurimpia soluelementtejä.
Näiden solujen kypsymisessä useita tiettyjä vaiheita erottuvat. Esimerkiksi useiden ytimien (Polyploidia) hankkiminen peräkkäisten solujen jakautumisten kautta, joissa DNA kerrotaan, mutta sytokiinia ei ole. DNA: n lisääntymisen lisäksi erityyppiset rakeet kertyvät myös.
Lähde: Wbensmith [CC 3: lla.0 (https: // creativecommons.Org/lisenssit/by/3.0)] Suurin osa näistä soluista sijaitsee luuytimessä, missä ne vastaavat alle 1% kokonaiskennoista. Tästä alhaisesta solun osuudesta huolimatta yhden kypsän megakariosyytin pirstoutuminen aiheuttaa monia verihiutaleita vuosina 2000 - 7000 verihiutaleita, prosessissa, joka kestää enemmän tai vähemmän viikossa.
Verihiutaleiden läpimenoa. Sarja molekyylielementtejä - lähinnä trombopoietiinia - on vastuussa prosessin järjestämisestä.
Näistä soluista johdetut elementit ovat verihiutaleita, joita kutsutaan myös trombosyyteiksi. Nämä ovat pieniä solujen solufragmentteja ja puuttuvat ydin. Verihiutaleiden havaitaan olevan osa verta ja ne ovat perustavanlaatuisia veren hyytymisprosessissa tai hemostaasissa, haavojen paranemisessa, angiogeneesissä, tulehduksessa ja luontaisessa immuniteetissa.
[TOC]
Historiallinen näkökulma
Prosessia, jolla verihiutaleet ovat peräisin, on tutkittu yli 100 vuotta. Vuonna 1869 Italian biologi nimeltään Giulio Bizzozero kuvasi, mikä näytti olevan jättiläinen solu, halkaisijaltaan yli 45 um.
Nämä erikoiset solut (niiden koon suhteen) eivät kuitenkaan liittyneet verihiutaleiden alkuperään vasta vuonna 1906. Tutkija James Homer Wright totesi, että alun perin kuvatut jättiläiskennot olivat verihiutaleiden edeltäjiä, ja kutsuivat megakasyytejä.
Myöhemmin mikroskopiatekniikoiden edistymisen myötä selvitettiin näiden solujen rakenteelliset ja toiminnalliset näkökohdat, joissa pika- ja brinkhous -osuudet korostetaan tälle kentälle.
Ominaisuudet ja rakenne
Megakariosyyttit: verihiutalevan vanhemmat
Megakariosyyttit ovat soluja, jotka osallistuvat verihiutaleiden syntymiseen. Kuten nimestä voi päätellä, megakariosyytti on suuri, ja sitä pidetään hematopoieettisten prosessien suurimpana soluna. Sen mitat ovat halkaisijaltaan 50–150 um.
Ydin ja sytoplasma
Korostuksensa lisäksi yksi tämän solulinjan näkyvimmistä ominaisuuksista on useiden ytimien läsnäolo. Kiinteistön ansiosta sitä pidetään polyploidisoluna, koska siinä on enemmän kuin kaksi kromosomipeliä näiden rakenteiden sisällä.
Voi palvella sinua: ensisijainen spermatosyyttiUseiden ytimien tuotanto tapahtuu megakariosyyttien muodostumisessa megakarioblastille, jossa ytimessä voidaan jakaa niin monta kertaa, että megakariosyyttissä on keskimäärin 8–64 ytimiä, keskimäärin. Nämä ytimet voivat olla hyperlobulaattia. Tätä tapahtuu endomitoosi -ilmiö, josta keskustellaan myöhemmin.
Megakaioraisyyttejä on kuitenkin myös ilmoitettu, että siinä on vain yksi tai kaksi ytimiä.
Sytoplasman suhteen se kasvaa merkittävästi sen tilavuuden suhteen, jota seuraa jokainen jakoprosessi ja esittelee suuren määrän rakeita.
Sijainti ja määrä
Näiden solujen tärkein sijainti on luuydin, vaikka niitä voidaan löytää myös vähemmässä määrin keuhkoissa ja pernassa. Normaaliolosuhteissa megakiaiosyytit vastaavat vähemmän kuin 1% kaikista johdon soluista.
Näiden progenitorisolujen huomattavan koon takia keho ei tuota suurta määrää megakariosyyttejä, koska yksi solu aiheuttaa monia verihiutaleita - toisin kuin muiden soluelementtien tuotanto, jotka tarvitsevat useita progenitorisoluja.
Keskimääräisessä ihmisessä voidaan muodostaa jopa 108 Megakariosyyttejä päivittäin, mikä aiheuttaa yli 10yksitoista verihiutaleet. Tämä verihiutaleiden määrä auttaa ylläpitämään kiertävän verihiutaleiden paikallaan olevaa tilaa.
Viimeaikaiset tutkimukset ovat korostaneet keuhkokudoksen merkitystä verihiutaleiden muodostavana alueena.
Funktiot
Megakariosyyttit ovat välttämättömiä soluja prosessille, jota kutsutaan trombopoyesiksi. Jälkimmäinen koostuu verihiutaleiden muodostumisesta, jotka ovat 2–4 um: n soluelementtejä, pyöristettyjä tai munanmuotoja, joilla ei ole ydinrakennetta ja sijaitsevat verisuonten sisällä verenkomponentteina.
Kuten ytimestä puuttuu, hematologit kutsuvat heitä solujen "fragmentteiksi" eikä soluiksi sellaisenaan - samoin kuin punaiset ja valkosolut.
Näillä solufragmenteilla on ratkaiseva rooli veren hyytymisessä, ylläpitää verisuonten eheyttä ja osallistuvat tulehduksellisiin prosesseihin.
Kun vartalo kokee jonkin tyyppistä haavaa, verihiutaleilla on kyky tarttua nopeasti toisiinsa, missä proteiinin eritys alkaa, joka alkaa hyytymän muodostumisen muodostumisen.
Koulutus ja kypsyminen
Koulutusjärjestelmä: Megakarioblastista verihiutaleille
Kuten edellä mainittiin, Megakaiorayte on yksi verihiutaleiden esiastesoluista. Kuten muiden soluelementtien synty, verihiutaleiden muodostuminen - ja siten megakariosyyttit - alkaa tavaratilan solulla (englannista Kantasolu) Multipotentiaalisilla ominaisuuksilla.
Megakarioblast
Prosessin solujen edeltäjät alkavat rakenteella, nimeltään Megacarioblast, joka kaksinkertaistaa sen ytimen, mutta ei kaksinkertaistaa täydellisen solun (tämä prosessi tunnetaan kirjallisuudessa nimellä Endomitoos) megakariosyytin muodostamiseksi.
Se voi palvella sinua: eukaryoottinen soluPromisecariocito
Vaihe, joka esiintyy heti megakarioblastien jälkeen, kutsutaan Promegacariocitoksi, sitten rakeinen megakariosyytti tulee ja lopulta verihiutale.
Ensimmäisessä tilassa solun ydin esittelee joitain lohkoja ja protoplasma on basofiilistä. Kun megakariosyyttivaihe lähestyy, protoplasma kääntyy asteittain eosinofiiliseksi.
Rakeinen megakariosyytti
Megakariosyyttien kypsymisessä liittyy menetys kyvyn lisääntymiselle.
Kuten nimi viittaa siihen.
Kun kypsä megakariosyytti on suunnattu Medulla -verisuonen sinimuotoisen endoteelisoluun ja käynnistää sen polun verihiutaleiden megakariosyyttinä
Verihiutale -megakariosyytti
Verihiutaleiksi kutsuttujen megakariosyyttien toiselle tyypiksi on ominaista digitaalisten laajennusten säteily, joka syntyy solukalvosta, nimeltään protoplasmisiksi herniaatiot. Näille alueille ovat edellä mainitut rakeet.
Solun kypsymisen edetessä jokainen herniaatio kärsii kuristumisesta. Tämän hajoamisprosessin tulos päättyy solufragmenttien vapauttamiseen, jotka ovat vain jo muodostettuja verihiutaleita. Tässä vaiheessa melkein koko megakariosyyttisytoplasma muuttuu pieniksi verihiutaleiksi.
Sääntelytekijät
Kuvattuja eri vaiheita, jotka vaihtelevat megakarioblastista verihiutaleisiin. Megakariosyytin kypsyminen on viivästynyt matkan varrella osteoblastisesta kapeasta verisuoniin.
Tämän reitin aikana kollageenikuiduilla on perustavanlaatuinen rooli Protoplaquet -muodostumisen estämisessä. Sitä vastoin verisuonen kapealla vastaava solumatriisi on runsaasti von willebrand- ja fibrinogeenitekijää, jotka stimuloivat trombopopousis.
Muita megakariosytopoyesin sääteleviä tekijöitä ovat muun muassa sytokiinit ja kasvutekijät, kuten trombopoietiini, interleupiinit. Trombopoietiinia todetaan erittäin tärkeänä säätelijänä koko prosessin ajan lisääntymisestä solujen kypsyyteen.
Lisäksi kun verihiutaleet kuolevat (ohjelmoitu solukuolema) ilmentävät kalvossa fosfatidilseriiniä poistoa edistämiseksi monosyytti-makrofagijärjestelmän ansiosta. Tämä solun ikääntymisprosessi liittyy glykoproteiinien deialinisointiin verihiutaleissa.
Jälkimmäiset tunnistavat maksasolujen Ashwell-Morell-nimiset reseptorit. Tämä edustaa ylimääräistä mekanismia verihiutaleiden poistamiseksi.
Tämä maksa -tapahtuma indusoi trombopoietiinisynteesin verihiutaleiden synteesin aloittamiseksi uudelleen, joten se toimii fysiologisena säätelijänä.
Voi palvella sinua: karioklesisEndomitoosi
Merkittävin - ja utelias.
Se koostuu DNA: n replikaatiosykleistä, jotka on irrotettu sytokiinien tai solujen jakautumisen kanssa sinänsä. Elinkaaren aikana solu kulkee 2N: n proliferatiivisen tilan läpi. Solun nimikkeistössä n käytetään haploidin nimeämiseen, 2N vastaa diploidi -organismia ja niin edelleen.
Tilan 2N jälkeen solu aloittaa endomitoosiprosessin ja alkaa asteittain kerätä geneettistä materiaalia, nimittäin: 4N, 8N, 16N, 64N ja niin edelleen. Joissakin soluissa geenikuormituksia on löydetty jopa 128N.
Vaikka tämän jakautumisen järjestäviä molekyylimekanismeja ei tiedetä tarkasti, tärkeä rooli johtuu epämuodostumien sytokineesituotteesta, jota löytyy myosiini II -proteiineissa ja aktiini F -aktiinissa.
Viitteet
- Alberts, b., Bray, D., Hopkin, k., Johnson, a. D -d., Lewis, J., Raff, m.,… & Walter, P. (2013). Välttämätön solubiologia. Garlantitiede.
- Alonso, m. -Lla. S., & I Pons, E. C. (2002). Kliinisen hematologian käytännön käsikirja. Antares.
- Arber, D. -Lla., Glader, b., Lista. F., Tarkoittaa, r. T., Paraskevas, f., & Rodgers, G. M. (2013). Wintroben kliininen hematologia. Lippinott Williams & Wilkins.
- Dacie, J. V., & Lewis, S. M. (1975). Käytännöllinen hematologia. Churchill Livingstone.
- Hoffman, R., Benz Jr, ja. J -., Silberstein, L. JA., Heslop, h., Anastasi, J., & Weitz, J. (2013). Hematologia: Perusperiaatteet ja käytännöt. Elsevier Health Sciences.
- Junqueira, L. C., Carneiro, J., & Kelley, R. JOMPIKUMPI. (2003). Perushistologia: Teksti ja atlas. McGraw-Hill.
- Kierszenbaum, a. Lens., & Kolme, l. (2015). Histologia ja solubiologia: Johdatus patologiaan e-kirja. Elsevier Health Sciences.
- Manascero, a. R -. (2003). Solujen morfologian atlas, muutokset ja niihin liittyvät sairaudet. KULMAKARVA.
- Marder, V. J -., Aird, W. C., Bennett, J. S., Schulman, S., & Valkoinen, G. C. (2012). Hemostaasi ja tromboosi: Perusperiaatteet ja kliininen käytäntö. Lippinott Williams & Wilkins.
- Nurden, a. T., Nurden, P., Sanchez, m., Andia, minä., & Anitua, ja. (2008). Paleletit ja haavan paraneminen. Biotieteen rajat: Lehti- ja virtuaalikirjasto, 13, 3532-3548.
- Pollard, t. D -d., Earnshaw, W. C., Lippincott-Schwartz, J., & Johnson, G. (2016). Solubiologia e-kirja. Elsevier Health Sciences.
- Rodak, b. F. (2005). Hematologia: perusteet ja kliiniset sovellukset. Ed. Pan -American Medical.
- San Miguel, J. F., & Sánchez-guijo, f. (Toim.-A. (2015). Hematologia. Perusteltu peruskäsikirja. Elsevier Espanja.
- Vives Corrons, J. Lens., & Aguilar Bascompte, J. Lens. (2006). Laboratoriotekniikan käsikirja hematologiassa. Masson.
- Welsch, u., & Sobotta, J. (2008). Histologia. Ed. Pan -American Medical.
- « Vilakarjojen ominaispiirteet, elinympäristö, viljely, hoito
- Vesililja -ominaisuudet, elinympäristö, hoito, käyttö »