

Proteiinit g rakenne, tyypit ja toiminnot
- 1486
- 179
- Eddie Hackett
Se G jompikumpi Guaniininukleotidia sitovat proteiinit, Ne ovat proteiineja, jotka liittyvät plasmamembraaniin, joka kuuluu signaalien "kytkentä", joilla on tärkeitä toimintoja monissa signaalinsiirtoprosesseissa eukaryoottisissa organismeissa.
Kirjallisuudessa G -proteiineja kuvataan binaarisina molekyylivaihtoina, koska niiden biologinen aktiivisuus määritetään niiden rakenteen muutoksilla, jotka nukleotidilajit ovat antaneet, joihin ne kykenevät liittymään: guanosiininukleotidit (difosfaatti (GDP) ja tyyposfaatti (GTP)).
RAS -proteiinin rakenne, monomeerinen G -proteiini (lähde: Mark 'AbSturz' [julkinen domeeni] Wikimedia Commonsin kautta) Ne aktivoivat yleensä GPCR -kytkettyjen reseptoreina tunnetun proteiiniperheen reseptorit (GPCR, englanti "G-proteiiniin kytketyt vastaanottimet "), jotka saavat ulkoisen alkuperäisen signaalin ja tekevät siitä konformaatiomuutokset, jotka laukaisevat aktivaation, joka myöhemmin kääntyy toisen efektoriproteiinin aktivointiin.
Jotkut kirjoittajat katsovat, että tämän proteiiniperheen koodaavat geenit kehittyivät esi -isien yhteisen geenin päällekkäisyydessä ja eroja, joiden tuotetta oli hienostunut ja erikoistunut yhä enemmän ja enemmän.
Näiden proteiinien monenlaisten solujen toimintojen joukossa ovat makromolekyylien translokaatio proteiinisynteesin aikana, hormonaalisten signaalien ja aistien ärsykkeiden transduktio sekä proliferaation ja solujen erilaistumisen säätely ja solujen erilaistuminen.
Tämän tyyppisestä proteiinista on kuvattu kaksi luokkaa: pienet G -proteiinit ja heterotrofimeeriset G -proteiinit. A G G -proteiinin kolme ensimmäistä dimensionaalista rakennetta saatiin yli vuosikymmenen pienestä GAS -tunnetusta G -proteiinista.
[TOC]
Rakenne
Rakenteellisesti ottaen tunnustetaan kaksi tyyppiä g -proteiineja: pienet G -proteiinit ja heterotrimic G -proteiinit, paljon monimutkaisempi.
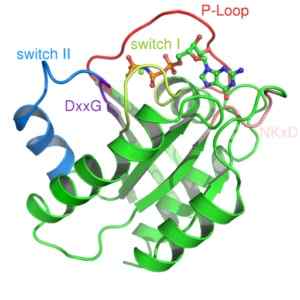
Pienet G-proteiinit muodostetaan yhdellä polypeptidillä, joka on noin 200 aminokeusa jätettä ja noin 20–40 kDa, ja rakenteessaan on säilytetty katalyyttinen domeeni (domeeni G), joka koostuu viidestä a-potkurista, kuudesta p-painettua arkkia ja viidestä polypeptidistä silmukot.
Toisaalta heterotrofimeeriset G -proteiinit ovat kattavia membraaniproteiineja, jotka koostuvat kolmesta polypeptidiketjusta, jotka tunnetaan nimellä a-, β- ja y -alayksiköt.
-A -alayksikkö painaa 40 - 52 kDa, sillä on guaniininukleotidien sitoutumisalue ja sillä on GTPASA -aktiivisuus hydrolysoiyhteyksiin GTP -fosfaattiryhmien välillä.
Eri G -proteiinien a -alayksiköillä on joitain rakenteellisia domeeneja, kuten GTP -liitos ja hydrolyysi, mutta ne ovat hyvin erilaisia reseptoreiden sitoutumiskohdissa ja efektoriproteiineissa.
Voi palvella sinua: mitoottinen kara-P -alayksikön molekyylipaino on hiukan alhaisempi (välillä 35 - 36 kDa).
-Γ -alayksikkö puolestaan on paljon pienempi ja sen likimääräinen molekyylipaino on 8 kDa.
Kaikki heterotropimeeriset G. Nämä kaksi verkkotunnusta liittyvät niin voimakkaasti, että niitä pidetään yhtenä funktionaalisena yksikkönä.
Kaverit
Kuten aikaisemmin mainittiin, G -proteiineja on kahta tyyppiä: pienet ja heterotrimaristiset.
Pienillä G -proteiineilla on toiminnot solujen kasvun, proteiinien erityksessä ja solunsisäisten rakkuloiden vuorovaikutuksessa. Heterotrofimeeriset G -proteiinit ovat puolestaan liittyviä signaalinsiirtoon pintareseptoreista ja toimivat myös kytkiminä, jotka vuorotellen kahden tilan välillä liittyvästä nukleotidista riippuen.
Pienet G -proteiinit
Näitä proteiineja kutsutaan myös pieniksi GTPASA: stä, pienistä GTP -liitoskäyttöproteiineista tai RAS -proteiinien superperheestä ja muodostavat riippumattoman superperheen suuressa hydrolaasi -GTP -luokassa säätelevien toimintojen kanssa.
Nämä proteiinit ovat hyvin monimuotoisia ja hallitsevat useita soluprosesseja. Niille on ominaista säilynyt domeeni, joka sitoutuu GTP: hen, domeeni "G". Tämän nukleotidifosfaatin liitto aiheuttaa pieniä konformaatiomuutoksia sen katalyyttisessä domeenissa GD -proteiineissa.
Sen aktiivisuus liittyy läheisesti GTPASA: iin, jotka aktivoivat proteiineja (aukko, englanti "GTPaasi aktivoi proteiinia ") ja guanina -nukleotidien vaihto -tekijöillä (GEF, englanti "Guaniinin nukleotidin vaihtotekijä ”).
Eukaryooteissa on kuvattu viisi luokkaa tai pienten G -proteiinien luokkaa tai perheitä:
-Rasva
-Rho
-Rab
-SAR1/ARF
-Juoksua
RAS- ja Rho -proteiinit kontrolloivat geneettistä ilmentymistä ja Rho -proteiinit myös moduloivat sytoskeleton uudelleenorganisointia. Rab- ja SAR1/ARF -ryhmäproteiinit ovat vaikutuksia vesikulaariseen kuljetukseen ja proteiinit säätelevät ydinkuljetusta ja solusykliä.
Heterotrofimeeriset G -proteiinit
Tämäntyyppinen proteiini ansaitsee myös yhteyden kahden muun proteiinitekijän kanssa siten, että signalointireitti ulkopuolelta solun sisäosaan koostuu kolmesta elementiä seuraavassa järjestyksessä:
- Se kytkettyjä reseptoreita A proteiinit g
- Se G
- Se Proteiinit tai Tehokas kanava
Heterotrofimeeristä G -proteiinia on suuri monimuotoisuus, ja tämä liittyy luonnossa esiintyvien a -alayksiköiden suureen monimuotoisuuteen, jossa vain 20% aminohapposekvenssistä säilyy.
Yleensä heterotrofimeeriset G -proteiinit tunnistetaan a -alayksikön monimuotoisuuden ansiosta, perustuen pääasiassa niiden funktionaalisiin ja sekvenssien samankaltaisuuksiin.
Α -alayksiköt muodostavat neljä perhettä (GS -perhe, GI/O -perhe, GQ -perhe ja G12 -perhe). Jokainen perhe koostuu erilaisesta ”isotyypistä”, joka yhdessä lisää yli 15 eri tapaa α -alayksiköitä.
Perhe Gs
Tämä perhe sisältää edustajia, jotka osallistuvat myös syklaasi -adenylaattiryhmien positiiviseen säätelyyn ja jotka ilmenevät useimmissa solutyypeissä. Se koostuu kahdesta jäsenestä: GS ja Golf.
Alaindeksi "S Se viittaa stimulaatioon (englannista "Stimulaatio ") ja alaindeksi "Olf " Se viittaa "hajuun" (englannista "Haju"-A. Golfproteiinit ekspressoivat erityisesti hajusta vastaavissa aistineuroneissa.
Perhe GI/O
Tämä on suurin ja monipuolinen perhe. Ne ekspressoivat monissa solutyypeissä ja välittävät monentyyppisten adenyylikassien vastaanottavasta riippuvaista estämistä (alaindeksi "Yo " viittaa estämiseen).
Proteiinit, joilla on ryhmä a.
Perhe GQ -
Proteiinit, joilla on tämän a -alayksikön perheen kanssa, ovat vastuussa fosfolipaasi C: n säätelystä. Tämä perhe koostuu neljästä jäsenestä, joiden α -alayksiköt ilmenevät eri geeneillä. Niitä on runsaasti maksasoluissa, munuaissoluissa ja keuhkoissa.
Perhe G12
Tämä perhe ekspressoituu kaikkialla organismeissa, eikä varmuudella tiedetä, mitä tarkalleen näiden alayksiköiden proteiinien avulla säätelee proteiinien kautta.
Β- ja y -alayksiköt
Vaikka alfa -rakenteiden monimuotoisuus on ratkaiseva heterotimeeristen proteiinien tunnistamisessa, kahdessa muussa alayksikössä on myös paljon monimuotoisuutta: beeta ja gamma.
Funktiot
G -proteiinit osallistuvat signaalien "kanavointiin" plasmamembraanin reseptoreista efektorikanaviin tai entsyymeihin.
Yleisin esimerkki tämäntyyppisen proteiinin toiminnasta on adenylaatti syklasa-entsyymin säätelyssä, entsyymissä, jotka vastaavat adenosiinin 3 ', 5'-monofosfaatin tai yksinkertaisesti syklisen AMP: n, molekyylin, jolla on tärkeät toiminnot toiseksi lähettiläänä, synteesistä. Monissa tunnetuissa soluprosesseissa:
Voi palvella sinua: kaliform -solut-Selektiivinen proteiinifosforylaatio spesifisillä toiminnoilla
-Geneettinen transkriptio
-Sytoskeleton uudelleenorganisointi
-Eritys
-Kalvon depolarisaatio
He osallistuvat myös epäsuorasti inositolien signaloimien vesiputouksen (fosfatidyylinitoli ja niiden fosforyloituneiden johdannaisten) säätelyyn, jotka ovat vastuussa kalsiumista riippuvien prosessien, kuten kemiotaxiksen, hallinnasta ja liukoisten tekijöiden erityksestä.
Monia ionisia kanavia ja kuljetusproteiineja kontrolloivat suoraan proteiiniproteiineilla G. Samoin nämä proteiinit ovat mukana monissa aistiprosesseissa, kuten visio, haju muun muassa.
Kuinka he toimivat?
G -proteiinin vuorovaikutusmuoto efektoriproteiineilla on tyypillinen jokaiselle luokalle tai proteiiniperheelle.
G-grebraniaalireseptori G-proteiineille (heterotrofimeeriset G-proteiinit) guaniininukleotidin, kuten BKT: n tai Guanosín-difosfaatin alayksikkö α kalvoon.
Jos BKT-molekyyli vaihdetaan myöhemmin GTP-molekyylillä, GTP: hen kiinnitetty a-alayksikkö dissosioituu β- ja γ-alayksiköistä, muodostaen erillisen kompleksin, joka tunnetaan nimellä Ga-GTP, joka pystyy muuttamaan sen entsyymien tai Diana-välittäjän aktiivisuutta proteiinit.
Tämän alayksikön hydrolyyttinen aktiivisuus antaa sinun lopettaa aktivoinnin, vaihtamalla GTP: n uudelle BKT: lle, siirtymällä passiiviseen konformaatioon.
Koska BKT: n BKT: n vaihtamisprosessiin liittyvät viritetyt reseptorit puuttuvat, se on erittäin hidas, mikä tarkoittaa, että heterotrimic GD -proteiinit vaihtavat BKT: n vain GTP: n avulla fysiologisesti tärkeällä nopeudella, kun he liittyvät heidän innostuneisiin reseptoreihinsa.
Viitteet
- Gilman, G. (1987). G-proteiinit: Vastaanottimen geneoitujen signaalien muuttajat. Vuotuiset arvostelut biokemiassa, 56, 615-649.
- Milligan, G., & Kostenis, ja. (2006). Heterotrimic g-proteiinit: lyhyt historia. British Journal of Farmacology, 147, 546-555.
- Offermanns, S. (2003). G-proteiinit transduereina kalvon läpäisevässä signaloinnissa. Biofysiikan ja molekyylibiologian edistyminen, 83, 101-130.
- Simon, m., Strathmann, M. P., & Gautam, N. (1991). G -proteiinien monimuotoisuus signaalinsiirrossa. Tiede, 252, 802-808.
- Syrovatkina, V., Iloinen, k. JOMPIKUMPI., Dey, r., & Huang, x. (2016). G-proteiinien säätely, signalointi ja fysiologiset toiminnot. Journal of Molecular Biology, 428(19), 3850-3868.
- « Crotalus triseriatus -ominaisuudet, elinympäristö, lisääntyminen
- Thunder -ominaisuudet, elinympäristö, taksonomia, kiinteistöt »