

Kalvoreseptorit toimivat, tyypit, miten ne toimivat
- 2018
- 18
- Shawn Stanton II
Se Membraanireseptorit Ne ovat tyyppisiä solureseptoreita, jotka sijaitsevat solujen plasmamembraanin pinnalla, jolloin ne voivat havaita kemiallisia aineita, jotka eivät luonteeltaan voi ylittää kalvoa.
Yleensä membraanireseptorit ovat kattavia membraaniproteiineja, jotka ovat erikoistuneet kemiallisten signaalien, kuten peptidihormonien, välittäjäaineiden ja tiettyjen troofisten tekijöiden havaitsemiseen; Jotkut lääkkeet ja toksiinit voivat myös liittyä tämän tyyppisiin vastaanottajiin.
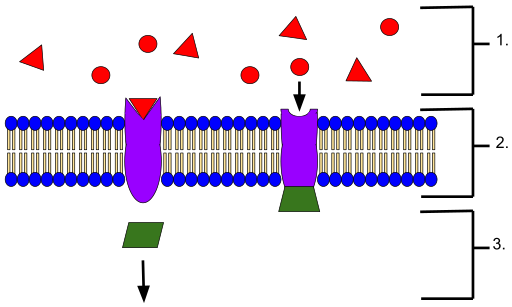
 Kalvovastaanottimen edustajajärjestelmä. Kalvon ulkopuolella (1) sijaitsevat ligandit, membraaniligandireseptorin vuorovaikutus (2) ja (3) seuraavat signalointitapahtumat (lähde: Wyatt Pyzynski [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta)
Kalvovastaanottimen edustajajärjestelmä. Kalvon ulkopuolella (1) sijaitsevat ligandit, membraaniligandireseptorin vuorovaikutus (2) ja (3) seuraavat signalointitapahtumat (lähde: Wyatt Pyzynski [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta) Ne luokitellaan solunsisäisen vesiputouksen tyypin mukaan, johon ne kytketään ja jotka määrittävät lopullisen vaikutuksen vastaavaan soluun, nimeltään kohdesolu tai valkoinen solu.
Siten on kuvattu kolme suurta ryhmää: ionisiin kanaviin liittyvät, entsyymeihin liittyvät ja G -proteiiniin kytketyt. Vastaanottimien ligandien liitto tuottaa konformaatiomuutoksen vastaanottimessa, joka laukaisee solunsisäisen signaloinnin vesiputouksen kohdesolussa.
Membraanireseptoreihin kytketty signalointiketjut sallivat signaalit monistaa ja tuottaa ohimeneviä tai pysyviä muutoksia kohdesoluissa. Näitä solunsisäisiä signaaleja kutsutaan yhdessä "signaalinsiirtojärjestelmään".
[TOC]
Funktiot
Membraanireseptoreiden ja muun tyyppisten reseptorien yleensä on sallia solujen viestintä toistensa kanssa siten, että organismin toiminnan eri elimet ja järjestelmät koordinoidulla tavalla ylläpitää homeostaasia ja reagoida vapaaehtoisiin ja automaattisiin tilauksiin Hermoston myöntämä.
Siten plasmamembraaniin vaikuttava kemiallinen signaali voi laukaista monien toimintojen monistetun modifikaation solun biokemiallisissa koneissa ja laukaisevat spesifisten vasteiden moninkertaisuuden.
Se voi palvella sinua: kadherinas: rakenne, ominaisuudet ja toiminnotSignaalin vahvistusjärjestelmän kautta yksi ärsyke (linkitys) pystyy tuottamaan välittömiä, epäsuoria, epäsuoria muutoksia ja pitkät muutokset, modifioimalla esimerkiksi joidenkin geenien ekspressiota, esimerkiksi kohdesolu.
Kaverit
Solukeskusreseptorit luokitellaan niiden sijainnin mukaan: Kalvoreseptorit (ne, jotka altistuvat solukalvossa) ja solunsisäiset reseptorit (jotka voivat olla sytoplasmisen tai ydin).
Kalvoreseptorit ovat kolme tyyppiä:
- Linkitetty ionikanaviin
- Entsyymeihin kytketty
- Proteiini G
Ionisiin kanaviin kytketyt kalvoreseptorit
Ne kutsutaan myös ionisiksi kanaviksi ligandin ovella, ne ovat yhdistelmäkalvoproteiineja 4–6 alayksiköiden välillä, jotka on koottu siten, että ne jättävät keskuskanavan tai huokosen, jonka läpi ionit kulkevat toiselta kalvolta toiselle.
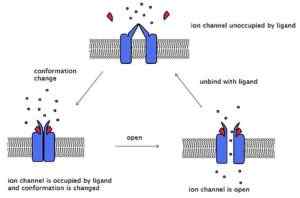 Esimerkki asetyylikoliinireseptorista, vastaanottimesta, joka on kytketty ioniseen kanavaan. Saman kolme konformaatiotilaa esitetään (lähde: Laozhengzz [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Wikimedia Commonsin kautta)
Esimerkki asetyylikoliinireseptorista, vastaanottimesta, joka on kytketty ioniseen kanavaan. Saman kolme konformaatiotilaa esitetään (lähde: Laozhengzz [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Wikimedia Commonsin kautta) Nämä kanavat ylittävät kalvon ja niillä on solunulkoinen pää, missä sitoutumispaikka sijaitsee, ja toinen solunsisäinen pää, joka joillakin kanavilla esittelee porttimekanismin. Tietyillä kanavilla on solunsisäinen ligandisivusto.
Entsyymi -sidotut membraanireseptorit
Nämä reseptorit ovat myös transmarkettiproteiineja. Heillä on solunulkoinen pää, joka esittelee ligandin unionin kohtaa ja jotka liittyvät heidän solunsisäiseen päätään entsyymi, joka aktivoituu ligandin liigan kanssa vastaanottimeen.
Kalvoreseptorit kytkettynä tai proteiini G
GFT -proteiinireseptoreilla on epäsuora mekanismi kohdesolujen solunsisäisten toimintojen säätelemiseksi, joka sisältää transduktiiviset molekyylit, joita kutsutaan GTP: n tai GTP: n tai proteiinien kiinnitysproteiineiksi.
Voi palvella sinua: Perusarkki: Ominaisuudet, histologia ja toiminnotKaikki nämä GP -proteiinireseptorit muodostuvat kalvoproteiinilla, joka ylittää kalvon seitsemän kertaa ja jota kutsutaan metabotrooppisiksi reseptoreiksi. Satoja reseptoreita, jotka on kytketty eri G -proteiineihin, on tunnistettu.
Kuinka he toimivat?
Ionisiin kanaviin kytketyissä reseptoreissa ligandin liitto vastaanottimeen tuottaa konformaatiomuutoksen vastaanottimen rakenteessa, joka voi muokata porttia, tuoda tai poistaa kanavan seinät. Tämän avulla he muokkaavat ionien kulkua kalvon toiselta puolelta.
Ionikanaviin kytketyt reseptorit ovat enimmäkseen spesifisiä ionityypille, joten K+-kanavien reseptoreita on kuvattu Cl-, Na+: sta, Ca ++: sta jne. On myös kanavia, jotka antavat kahden tai useamman ionin läpikulun.
Suurin osa entsyymiin liittyvistä reseptoreista liittyy kinaasiproteiineihin, etenkin kinaasityrosiinientsyymillä. Nämä kinaasit aktivoituvat, kun reseptorin sitoutuminen sen solunulkoiseen unionin kohtaan. Quinaas fosforyloi spesifiset proteiinit kohdesolussa, joka muuttaa saman toimintaa.
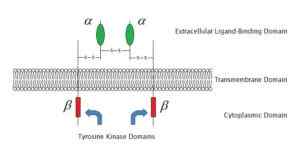 Esimerkki membraanireseptorista, joka on kytketty entsyymityrosiinikinaasiin (lähde: Laozhengzz [julkinen alue] Wikimedia Commons -sovelluksen kautta)
Esimerkki membraanireseptorista, joka on kytketty entsyymityrosiinikinaasiin (lähde: Laozhengzz [julkinen alue] Wikimedia Commons -sovelluksen kautta) Proteiiniabinked reseptorit -aktiviteetti.
G -proteiineja on kahta tyyppiä, jotka ovat heterotrimic g -proteiineja ja monomeerisiä G -proteiineja. Molemmat on kytketty BKT: hen epäaktiivisesti, mutta kun vastaanottimen ligandi korvataan BKT: llä GTP: llä ja G -proteiini aktivoidaan.
Heterotrimic G -proteiineissa a -alalause, joka on kytketty GTP: hen dissosiaatiota ßy -kompleksista, jättäen proteiini G: n aktivoitumaan. Sekä GTP: hen kytketty a -alayksikkö että vapaa ßy voi välittää vastausta.
Voi palvella sinua: trofoblast GF -proteiinin kytkettyjen vastaanottimen kaavio (lähde: Bensacount englanniksi Wikipedia [julkinen alue] Wikimedia Commonsin kautta)
GF -proteiinin kytkettyjen vastaanottimen kaavio (lähde: Bensacount englanniksi Wikipedia [julkinen alue] Wikimedia Commonsin kautta) Monomeerinen G -proteiinit tai pienet G -proteiinit saavat myös RAS -proteiinien nimen, jota kuvataan ensimmäistä kertaa viruksessa, joka tuottaa Sarcommon -kasvaimia rotilla.
Aktivoituna ne stimuloivat mekanismeja, jotka liittyvät pääasiassa vesikulaariseen liikenteeseen ja sytoskeleton funktioihin (modifikaatio, uusinta, kuljetus jne.-A.
Esimerkit
Asetyylikoliinireseptori, joka on kytketty natriumkanavaan, joka aukeaa asetyylikoliinia ja tuottaa kohdesolujen depolarisaation, on hyvä esimerkki ionisiin kytkettyihin kalvoreseptoreihin, jotka on kytketty ionisiin. Lisäksi on kolmen tyyppisiä glutamaattireseptoreita, jotka ovat ionotrooppisia reseptoreita.
Glutamaatti on yksi hermoston tärkeimmistä eknitutiivisista neurotransmittereista. Sen kolme ionotrooppisia reseptoreita ovat: NMDA (N-metyyli-D-spartaatti), AMPA (α-amino-3-hydroksi-5-metyyli-4-isoksatsoli-propionaatti) ja kainato) ja kainato (happo kainico).
Heidän nimensä ovat peräisin agonisteista, jotka aktivoivat niitä, ja nämä kolme kanavaa ovat esimerkkejä ainutlaatuisista jännittävistä kanavista, koska ne sallivat natriumin ja kaliumin kulkemisen ja joissain tapauksissa pieniä määriä kalsiumia.
Esimerkkejä entsyymistä linkitetyistä reseptoreista ovat insuliinireseptori, TRK -reseptorien perhe tai neurotrofiinireseptorit ja reseptorit joillekin kasvutekijöille.
Tärkeimpiä proteiini-G-reseptoreita ovat asetyylikoliinin muskariinireseptorit, β-adrenergiset reseptorit, hajujärjestelmän reseptorit, metabotrooppiset glutamaattireseptorit, monien peptidihormonien reseptorit ja verkkokalvon järjestelmän rodopsiinireseptorit.
Viitteet
- Biokemian ja molekyylin biofysiikan laitos Thomas Jesell, Siegelbaum, S., & Hudspeth, a. J -. (2000). Neuraalitieteen periaatteet (Vol. 4, pp. 1227-1246). JA. R -. Kandel, J. H. Schwartz, & t. M. Jesell (toim.-A. New York: McGraw-Hill.
- Hulme, E. C., Birdsall, n. J -. M., & Buckley, N. J -. (1990). Muskariinin vastaanottimen alatyypit. Farmakologian ja toksikologian vuosikatsaus, 30(1), 633-673.
- Cull-Candy, S. G., & Leszkiewicz, D. N. (2004). Erillisten NMDA -vastaanottimen alatyyppien rooli keskussynapsissa. Sci. Stelli, 2004(255), RE16-RE16.
- William, f. G., & Ganong, M. D -d. (2005). Katsaus lääketieteelliseen fysiologiaan. Painettu Amerikan yhdysvalloissa, seitsemäs painos, pp-781.
- Karhu, m. F., Connors, b. W -., & Paradiso, M. -Lla. (Toim.-A. (2007). Neurotiede (Vol. 2). Lippinott Williams & Wilkins.
- « Achlostoma caninum -ominaisuudet, morfologia, oireet
- Canelo -ominaisuudet, elinympäristö, ominaisuudet, viljely »