

DNA n replikaatiomekanismit prokaryooteissa ja eukaryooteissa
- 2975
- 238
- Louis Moen
Se Toisto jstk DNA (Desoksihiobonukleiinihappo) Se koostuu genomin kopioinnista, toisin sanoen kaikki organismin DNA: n geneettinen tieto, joka tuottaa kaksi identtistä kopiota. Genomilla on tarvittavat tiedot täydellisen organismin rakentamiseksi.
Ennen solun jakautumista DNA: n replikaatio tapahtuu. Meioosin kautta tapahtuu sukupuolisen lisääntymisen sukusolut. Mitoosin kautta solujen korvaaminen tapahtuu (esimerkiksi iho ja veri) ja kehitys (esimerkiksi kudokset ja elimet).
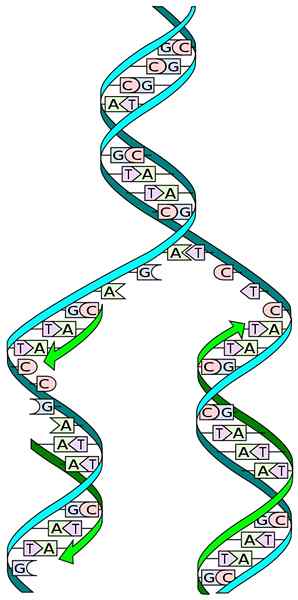
 Lähde: Minä, MadPrime [CC0]
Lähde: Minä, MadPrime [CC0] DNA -rakenteen tuntemus antaa meille ymmärtää tapaa, jolla sen replikaatio tapahtuu. DNA: n rakenne koostuu kaksoispotkurista, joka koostuu kahdesta peräkkäisten nukleotidien antipaallaalisesta ketjusta, joiden typpialusta on täydennetty erityisesti.
Replikaation aikana jokainen kaksois -DNA -ketjun ketjut toimivat muotina uuden ketjun biosynteesille. Kahdessa äskettäin syntetisoidussa ketjussa on emäksiä, jotka ovat täydentäviä muotiketjun emäksiä: adeniini (a) Timina (t) ja sytosiini (c) guaniinin (g) kanssa (g).
Useat entsyymit ja proteiinit osallistuvat DNA: n replikaatioon. Esimerkiksi kaksois-DNA-potkurin avaaminen, DNA: n pitäminen auki ja lisäämällä deoksihiobonukleosidos-5'-trifosfaattia (DNTP) uuden ketjun muodostamiseksi.
[TOC]
DNA: n replikaatio on puolikonservatiivinen
DNA: n rakenteen perusteella Watson ja Crick ehdottivat, että DNA: n replikaatio tapahtuu puolikonservatiivisella tavalla. Tämän osoittivat Meselson ja Stahl merkitsemällä DNA: ta Escherichia coli Typen raskas isotooppi, viisitoistaN, useiden sukupolvien seurauksena jakautumismalli viljelyalustalla kevyellä typellä, 14N.
Messelson ja Stahl havaitsivat, että ensimmäisessä sukupolvessa kaksi tytärtä DNA -molekyyliä oli, kukin molekyyli merkitty ketjulla raskaan typen isotooppin kanssa ja toinen kevyellä isotoopilla. Toisin kuin vanhempien DNA -molekyyli, jolla oli kaksi ketjua, jotka oli merkitty raskaalla isotoopilla, viisitoistaN.
Toisessa sukupolvessa 50% DNA -molekyyleistä oli kuin ensimmäisen sukupolven, ja muilla 50%: lla oli vain kevyt typpi. Tämän tuloksen tulkinta on, että kaksoistytärpotkurilla on vanhempien ketju (joka toimii muotina) ja uusi ketju.
Puoli -konservatiivinen replikaatiomekanismi merkitsee DNA -ketjujen erottamista ja täydentävien emäksen pariutumista peräkkäisten nukleotidien avulla, jotka tuottavat kaksi kaksinkertaista tytärtä tyttäriä tyttäriä.
Replikointi paristoissa
DNA: n replikaation aloittaminen bakteereissa
Bakteerien DNA koostuu pyöreästä kromosomista, ja siinä on vain yksi replikaation lähtökohta. Tästä sivustosta kahden tyttären biosynteesi tapahtuu kaksisuuntaisesti muodostaen kaksi replikointihaarukkaa, jotka liikkuvat lähtökohtana vastakohtana. Loppujen lopuksi haarukat ovat toistamalla replikaatio.
Replikaatio alkaa DNAA -proteiinien liitoksella lähtökohtaan. Nämä proteiinit puolestaan muodostavat kompleksin. Sitten Hu- ja IHF -proteiinit sitovat.
Seuraavaksi liittyy DNAC -proteiineja, jotka tekevät DNA -yhdistyksen helikaasista. Nämä auttavat rentoutumaan DNA: n ja rikkoutuvat vety sidokset, jotka on muodostettu emäspariin. Joten kaksi ketjua erottuvat vielä enemmän, muodostaen kaksi yksinkertaista ketjua.
Voi palvella sinua: mikä on kromosomaalinen permutaatio?Topoisomeraasi II tai DNA Girasa liikkuu DNA -helikyksen edessä. Yhden ketjun DNA: n sitoutumisproteiinit pitävät DNA -ketjut erillään. Siten tytärketjun biosynteesi voi alkaa.
Tyttäreiden biosynteesi DNA -ketjuissa bakteereissa
Prima -entsyymi on vastuussa lyhyiden RNA -ketjujen syntetisoinnista, joita kutsutaan alukkeiksi, joiden pituus on 10-15 nukleotidia. DNA-polymeraasi alkaa lisätä 5'-trifosfaattia (DNTPS) alustavan sokerin 3'-OH: hon, minkä jälkeen ketju kasvaa edelleen samassa päässä.
Koska DNA -ketjut ovat antiparallelia, pohjamaali syntetisoidaan opasketjussa ja monet viivästyneessä ketjussa olevat alukkeet. Tämän vuoksi viivästyneen ketjun biosynteesi on epäjatkuva. Vaikka DNA -ketjut ovat antiparallelia, replikaatiohaarukka liikkuu yhteen suuntaan.
DNA -polymeraasi on vastuussa äskettäin syntetisoitujen ketjujen vierekkäisten nukleotidien kovalenttisten sidosten muodostumisesta 5'®3 '-suunnassa. Sisään JA. koli, Polymeraasi -DNA: ta on viisi: DNA -polymeraasit I ja III suorittavat DNA: n replikaation; ja DNA -polymeraasit II, IV ja V ovat vastuussa vaurioituneen DNA: n replikaatiosta ja toistamisesta.
Suurin osa replikaatiosta tehdään DNA -polymeraasi III: lla, joka on holoentsyymi, jolla on 10 erilaista alayksikköä, joilla on useita toimintoja DNA: n replikaatiossa. Esimerkiksi Alpha -alayksikkö vastaa sidosten tekemisestä nukleotidien välillä.
Entsyymikompleksi on vastuussa DNA: n replikaatiosta bakteereissa
DNA: n ja Priman helikaasi yhdistetään kompleksin muodostamiseksi nimeltä Primosoma. Tämä liikkuu DNA: ta pitkin ja toimii koordinoidulla tavalla erottaa kaksi vanhempainketju.
Primosomi sitoutuu fyysisesti DNA -polymeraasi III: iin ja muodostaa replisomin. Kaksi DNA -polymeraasia III ovat vastuussa oppaan DNA: n replikaatiosta ja viivästyneistä ketjuista. DNA -polymeraasi III: n suhteen viivästynyt ketju muodostaa silmukan, mikä mahdollistaa nukleotidien lisäämisen tähän ketjuun samaan ohjausketjun suuntaan.
Nukleotidien lisääminen ohjausketjuun on jatkuvaa. Viivästyksessä se on epäjatkuva. Fragmentit, joissa on 150 pituuden nukleotidia, nimeltään Okazaki -fragmentit.
DNA -polymeraasin I eksonukleaasiaktiivisuus 5 ' -> 3' on vastuussa alukkeiden eliminoinnista ja täyttämisestä, lisäämällä nukleotideja. Entsyymiligaasi tiivistää fragmenttien väliset aukot. Replikointi päättyy, kun kaksi replikaatiota hoquillas ovat valmistumisjärjestyksessä.
Proteiini sitoutuu lopetussekvenssiin, pysäyttäen replikaatiohaarukan liikkumisen. Topoisomeraasi II mahdollistaa kahden kromosomin erottamisen.
Disoksibonukleotidit Tryfosfaatteja käyttää DNA -polymeraasi
Dexinukleosidi -tryfosfaatti (DNTP) sisältää kolme fosfaattiryhmää, jotka on yhdistetty hiili 5 ': n deoksihiobosaan. DNTPS (DATP, DTTP, DGTP ja DCTP) liittyy muottiketjuun AT/GC -säännön mukaisesti.
Voi palvella sinua: PlesiomorphyDNA-polymeraasi katalysoi seuraavaa reaktiota: kasvavan ketjun nukleotidin hydroksyyliryhmä (-OH) 3 'reagoi tulevan DNTP: n alfafosfaatin kanssa vapauttaen epäorgaanisen pyrofosfaatin (PPI) (PPI). PPI -hydrolyysi tuottaa energiaa kovalenttisen sidoksen tai fosfodiéster -sidoksen muodostumiseen kasvavan ketjun nukleotidien välillä.
Mekanismit, jotka varmistavat DNA: n replikaation uskollisuuden
DNA: n replikaation aikana DNA -polymeraasi III tekee virheen 100 miljoonaa nukleotideja kohti. Vaikka virheen todennäköisyys on erittäin alhainen, on mekanismeja, jotka varmistavat uskollisuuden DNA: n replikaatiossa. Nämä mekanismit ovat:
1) Stabiilisuus perusaparissa. Vety sidosenergia AT/GC: n välillä on suurempi kuin virheellisissä emäsparissa.
2) DNA -polymeraasin aktiivisen kohdan rakenne. DNA -polymeraasi katalysoi mieluiten nukleotideja oikealla emäksellä vastakkaisessa ketjussa. Päädoitusten huono pariutuminen aiheuttaa kaksinkertaisen DNA -potkurin vääristymisen, mikä estää väärän nukleotidin miehittämästä entsyymin aktiivista kohtaa.
3) Lukematesti. DNA -polymeraasi tunnistaa sisällytetyt virheelliset nukleotidit ja eliminoi ne tytärketjusta. Polymeraasi -DNA: n eksonukleaasiaktiivisuus katkaisee fosfodiéster -sidokset nukleotidien välillä uuden ketjun 3' -päässä.
DNA: n replikaatio eukaryooteissa
Toisin kuin prokaryootien replikaatio, jonka replikaatio alkaa yhdessä paikassa, eukaryoottinen replikaatio alkaa useista lähtökohdista ja replikaatiohaarukka siirtää kaksisuuntaisesti kaksisuuntaisesti. Myöhemmin kaikki replikaatiohaarukat yhdistetään, muodostaen kaksi sisarkromatidia, jotka on liitetty sentromeriin.
Eukaryotasilla on monen tyyppisiä DNA -polymeraasia, joiden nimet käyttävät kreikkalaisia kirjaimia. DNA -polymeraasi α muodostaa kompleksin prima. Tämä kompleksi syntetisoi lyhyet alukkeet, jotka koostuvat 10 RNA -nukleotidista, jota seuraa 20 - 30 DNA -nukleotidia.
Sitten DNA -polymeraasi ε jompikumpi Δ katalysoi tytärketjun pidentymistä pohjamaalista. DNA -polymeraasi ε Se osallistuu johtavan ketjun synteesiin, kun taas DNA -polymeraasi Δ Syntetisoi viivästynyt ketju.
DNA -polymeraasi Δ Pidennä vasenta Okazaki -fragmenttiä, kunnes saavutat oikean RNA -alukkeen, tuottaen lyhyen noston alukkeesta ("lyhyt läppä"). Toisin kuin prokaryootit, joissa polymeraasi -DNA eliminoi aluke.
Seuraavaksi DNA -ligaasi tiivistää vierekkäiset DNA -fragmentit. Replikaation loppuun saattaminen tapahtuu replikaatiohaarukkaproteiinien dissosiaatiolla.
Se DNA: n replikaatio eukaryooteissa ja solusyklissä
Replikaatio eukaryooteissa tapahtuu solusyklin S -vaiheessa. Replikoituneet DNA -molekyylit erotetaan kahteen tytärsoluun mitoosin aikana. Vaiheet G1 ja G2 erottavat S -vaiheen ja mitoosi. Kinaasit, fosfataasit ja proteaasit säätelevät voimakkaasti solusyklin kunkin vaiheen etenemistä.
Solusyklin G1 -vaiheessa alkuperätunnistuskompleksi (OCR) liittyy lähtökohtaan. Tämä indusoi MCM-helikasten ja muiden proteiinien, kuten CDC6: n ja CDT1: n, liiton ennalta replikaatiokompleksin (PRERC) muodostamiseksi (PRERC). Las Helicase MCM liittyy ohjausketjuun.
Voi palvella sinua: linkitetyt geenitVaiheessa S, PRERC: stä tulee aktiivinen replikointipaikka. OCR-, CDC6- ja CDT1 -proteiinit vapautuvat, ja MCM -helikaasi liikkuu suuntaan 3 ' - 5'. Kun replikaatio päättyy, tämä käynnistetään uudelleen seuraavassa solusyklissä.
Kromosomien päiden replikaatio eukaryooteissa
Kromosomien päät tunnetaan telomeereinä, jotka koostuvat toistuvista sekvensseistä samanaikaisessa.
DNA -polymeraasi ei pysty toistamaan DNA -ketjujen 3 'päätä. Tämä johtuu siitä, että DNA-polymeraasi voi syntetisoida DNA: ta vain 5'-3 '-osoitteessa, ja se voi pidentää vain olemassa olevia ketjuja, kykenemättä syntetisoimaan pohjamaalin tällä alueella. Tämän seurauksena telomeerejä lyhennetään jokaisessa replikaatiossa.
Telomeraasientsyymi estää telomeerejä lyhenemällä. Telomeraasi on entsyymi, jolla on proteiini- ja RNA -alayksiköitä (kolmas). Jälkimmäinen sitoutuu DNA: n toistuviin sekvensseihin ja antaa telomeraasin liittyä telomeerin 3' -päähän.
Unionin takana oleva RNA -sekvenssi, joka sijaitsee muotina kuuden nukleotidin (polymeroinnin) sekvenssin synteesille DNA -ketjun lopussa. Telomeerin pidennys katalysoi telomeraasin alayksiköt, joita kutsutaan käänteisen telomeraasin transkriptaasi (TERT).
Polymeroinnin jälkeen tapahtuu translokaalisaatio, joka koostuu telomeraasin liikkumisesta DNA -ketjun uuteen päähän, yhdistämällä kuusi muuta nukleotidia loppuun saakka.
Muiden DNA -polymeraasien toiminnot eukaryooteissa
DNA -polymeraasi β Sillä on tärkeä rooli väärien DNA: n emäksen poistamisessa, mutta se ei ole mukana DNA: n replikaatiossa.
Monet havaitsivat polymeraasi-DNA: n kuuluvan ”translesion repliscating” -polymeraasiryhmään. Nämä polymeraasit ovat vastuussa täydentävien ketjujen syntetisoinnista vaurioituneella DNA -alueella.
Polymeraaseja on useita tyyppejä "translesion replikoivia". Esimerkiksi DNA -polymeraasi η Voit toistaa Timina Dímerosista, jotka tuottavat UV -valo.
DNA: n replikaatio arkeobakteereissa
Arkeobakteerien DNA: n replikaatio on samanlainen kuin eukaryooteissa annettu. Tämä johtuu seuraavista: 1) replikaatioon osallistuvat proteiinit ovat enemmän kuin eukaryootien kuin prokaryootien proteiineja; ja 2) Vaikka replikaatiopaikka on vain yksi, kuten prokaryooteissa, sen sekvenssi on samanlainen kuin eukaryoottisen alkuperän paikka.
Kaarien ja eukaryootien välisen replikaation samankaltaisuus tukee ajatusta, että molemmat ryhmät ovat fylogeneettisesti enemmän suhteessa toisiinsa kuin mikään niistä prokaryootien kanssa.
Viitteet
- Brooker, r. J -. 2018. Genetiikan analyysi ja periaatteet. McGraw-Hill, New York.
- Hartwell, L. H., Goldberg, m. Lens., Fischer, J. -Lla., Luuku, l. 2018. Genetiikka - genomien geeneistä. McGraw-Hill, New York.
- Kušić-tišma, J. 2011. DNA: n replikaation perustavanlaatuiset näkökohdat. Intech Open Access, Kroatia.
- Lewis, r., 2015. Ihmisen genetiikan käsitteet ja sovellukset. McGraw-Hill, New York.
- Pierce, b. -Lla. 2005. Genetiikka - käsitteellinen lähestymistapa. W -. H. Freeman, New York.