

Succinato -dehydrogenaasirakenne, toiminta, säätely, sairaudet
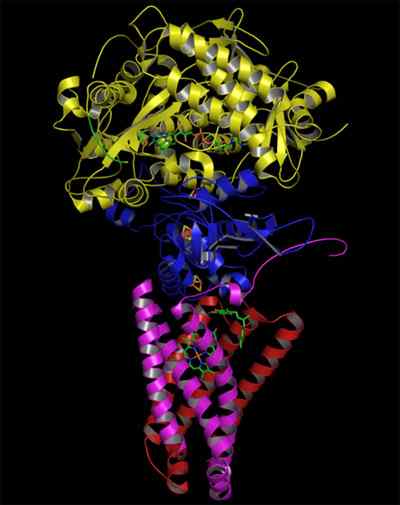
- 2766
- 88
- Sheldon Kuhn
SUccinato -dehydrogenaasi (SDH), joka tunnetaan myös nimellä elektronikuljetusketjun kompleksi II, se on mitokondriaalinen proteiinikompleksi, jolla on entsymaattinen aktiivisuus, joka toimii sekä Krebs -syklissä että elektronikuljetinketjussa (solujen hengitys).
Se on entsyymi, jota on läsnä kaikissa aerobisissa soluissa. Eukaryooteissa se on monimutkainen, joka liittyy läheisesti sisäiseen mitokondriaaliseen kalvoon, kun taas prokaryooteissa sitä löytyy plasmamembraanista.
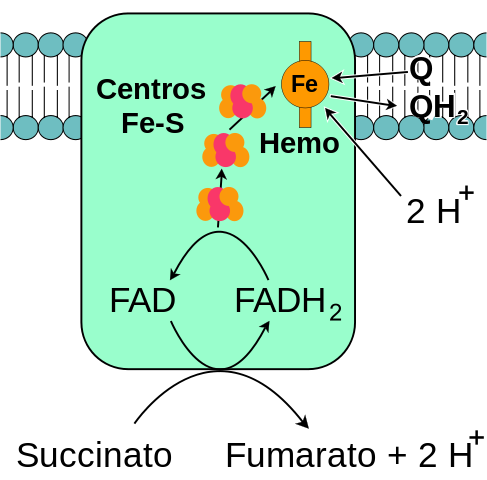 Mitokondriaalikompleksin sukkinaatti -dehydrogenaasin yleinen kaavio (lähde: minä, FVASCONCELLOS -vektorisoinnin perusteella. / Julkinen verkkotunnus, Wikimedia Commons) kautta)
Mitokondriaalikompleksin sukkinaatti -dehydrogenaasin yleinen kaavio (lähde: minä, FVASCONCELLOS -vektorisoinnin perusteella. / Julkinen verkkotunnus, Wikimedia Commons) kautta) Singer ja Kearney on tutkittu laajasti monista syistä monimutkaisia sukkinaattidehydrogenaasia, jota löydettiin noin vuonna 1910 ja puhdistettu ensimmäistä kertaa vuonna 1954:
- Se toimii sekä KREBS -syklissä (sitruunahapposykli tai trikarboksyylihapposykli) että elektronien kuljetusketjussa (katalysoi sukkinaatin hapettumista fumaraattiin)
- Sen aktiivisuutta säätelevät eri aktivaattorit ja estäjät ja
- Se on kompleksi, johon liittyy: rauta, jota ei liitetä Flavina Adeninan (FAD) hemo -ryhmään, labyylirikiin ja dyukleotideihin
Ydingenomi koodaa sen ja on osoitettu, että neljässä geenissä olevat mutaatiot, jotka kodivat sen alayksiköt (a, b, c ja d), johtavat erilaisiin kliinisiin kuviin, ts. Ne voivat olla melko negatiivisia pisteestä. ihmisten fyysinen koskemattomuus.
[TOC]
Rakenne
Ydingenomin koodaava entsyymikompleksi sukkinaattidehydrogenaasi muodostuu neljästä alayksiköstä (heterotarámero), joten se on ainoa oksidatiivisen fosforylaation kompleksi elektronikuljetinketjussa, jolla ei ole mitokondriogenomin koodaavaa alayksikköä.
Lisäksi tämä kompleksi on ainoa, joka ei pumppaa protoneja sisäisen mitokondriaalikalvon läpi katalyyttisen vaikutuksen aikana.
Sikien sydänsolujen entsymaattisen kompleksin perusteella olevien tutkimusten mukaan kompleksi sukkinaattidehydrogenaasi koostuu:
- a "pää" Hydrofiilinen joka ulottuu mitokondrioiden sisäisestä kalvosta mitokondriaaliseen matriisiin ja
- a "linja" Hydrofobinen joka on upotettu sisäiseen mitokondriaaliseen kalvoon ja jolla on pieni segmentti, joka projisoidaan mitokondrioiden liukoiseen memembraanialueeseen
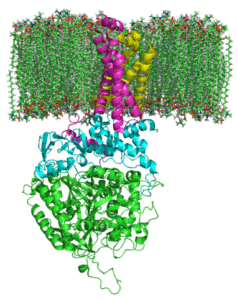 Kompleksin sukkinaatti-dehydrogenaasin rakenne (lähde: zephyris englannin kielellä Wikipedia/CC BY-SA (http: // creativecommons.Org/lisenssit/by-SA/3.0/) Wikimedia Commonsin kautta)
Kompleksin sukkinaatti-dehydrogenaasin rakenne (lähde: zephyris englannin kielellä Wikipedia/CC BY-SA (http: // creativecommons.Org/lisenssit/by-SA/3.0/) Wikimedia Commonsin kautta) Hydrofiilinen annosrakenne
Hydrofiilinen pää koostuu SDHA: sta (70 kDa) ja SDHB: stä (27 kDa) (SDH1 ja SDH2 hiivoissa) ja tämä käsittää kompleksin katalyyttisen keskuksen.
SDHA: n ja SDHB.
SDHA: n alayksikössä on villityskofaktori (koentsyymi, joka osallistuu oksidin vähentämisreaktioihin) yhdessä sen rakenteen kanssa, vain sukkinaatin risteyksen kohdalla (entsyymin pää substraatti).
SDHB. Yksi keskuksista, 2FE-2: sta, on lähellä SDHA-alayksikön ja muiden (4FE-4 ja 3FE-4) villityskohtaa ovat ensimmäisen vieressä.
Voi palvella sinua: fylogenyOn huomattava, että rakennetutkimukset osoittavat, että SDHB.
Hydrofobinen annosrakenne
Kompleksin kalvodomeeni, kuten todettiin, koostuu SDHC: stä (15 kDa) ja SDHD: stä (12-13 kDa) (SDH3 ja SDH4 hiivassa), jotka ovat kattavia kalvoproteiineja, jotka on muodostettu, 3 trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-trans-potilaiden avulla.
Tämä verkkotunnus sisältää hemo -osan b - Yhdistettynä SDHC: n ja SDHD.
Tässä entsyymissä on havaittu kaksi Ubiquinonan unionin sivustoa: yksi suuresta affiniteetista ja yksi matala affiniteetista.
Korkea affiniteettipaikka, joka tunnetaan nimellä QP (P mukaan proksimaalinen) Se on mitokondriaalimatriisin edessä ja muodostuu spesifisillä aminohappotähteillä, jotka sijaitsevat SDHB-, SDHC- ja SDHD.
Matala affiniteettipaikka, jota kutsutaan myös Qd (D kirjoittanut distaalinen) Se on sisäisen mitokondriaalisen kalvon osassa, johon kompleksi asetetaan lähemmäksi membraanien välistä tilaa, toisin sanoen organelle -matriisista.
Yhdessä kokonaiskompleksin molekyylipaino on lähellä 200 kDa ja on määritetty, että sen suhde on 4.2-5.0 flavin-nanomolit milligrammaa proteiinia kohti ja 2-4 g rautaa jokaiselle Flavinina-moolille.
Funktio
Entsymaattinen sukkinaattikompleksi dehydrogenaasi täyttää tärkeän toiminnon mitokondrioissa, koska ei vain osallistu Krebs -sykliin (missä se osallistuu asetyyli -CoA: n hajoamiseen), mutta myös osa hengitysketjua, joka on energiantuotannon perusta, ATP -muotoinen.
Toisin sanoen, se on keskeinen entsyymi välittäjäaineenvaihduntaan ja ATP: n aerobiseen tuotantoon.
- Se on vastuussa sukkinaatin hapettumisesta fumaraattiin sitruunahapposyklissä
- Se ruokkii elektronikuljetinketjun kompleksia III sukkinaatin hapettumisesta johdettujen elektronien kanssa, mikä auttaa vähentämään happea ja muodostavat vettä
- Elektronikuljetus tuottaa sähkökemiallisen gradientin sisäisen mitokondriaalisen kalvon läpi, joka suosii ATP -synteesiä
Vaihtoehtona elektroneja voidaan käyttää vähentämään ubikinonien ”uima -altaalla” olevia molekyylejä, mikä tuottaa tarvittavat pelkistimet vähentämään superoksidianioneja, jotka ovat peräisin samasta hengitysketjusta tai jotka tulevat eksogeenisistä lähteistä.
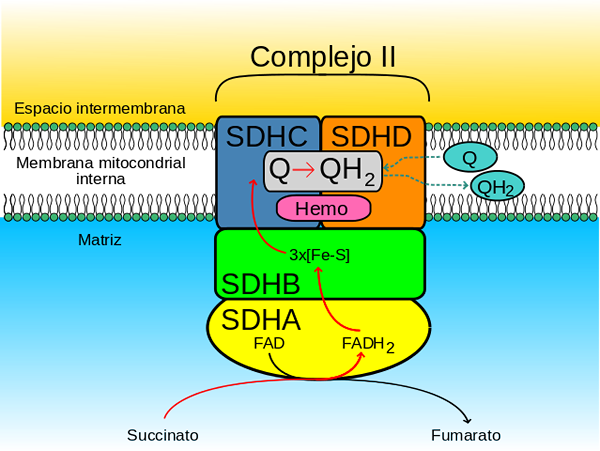 Sukkinaattikompleksi dugidrogenasa (lähde: Johnhfst / Public Domain, Wikimedia Commonsin kautta)
Sukkinaattikompleksi dugidrogenasa (lähde: Johnhfst / Public Domain, Wikimedia Commonsin kautta) Kuinka se toimii?
Kompleksin alayksikkö A (joka on kovalenttisesti kytketty fad -koentsyymiin) liittyy substraatteihin, fumaraattiin ja sukkinaattiin, samoin kuin sen fysiologisiin säätelijöihin, oksalaseettiin (kilpailukykyinen estäjä) ja ATP.
ATP syrjäyttää oksalasetaatin ja SDH -kompleksin välisen liiton ja silloin SDHA -alayksikön sukkinaatista "menneisyydet" siirretään SDHB.
Voi palvella sinua: myosiini: Ominaisuudet, rakenne, tyypit ja toiminnotAlayksiköstä B nämä elektronit saavuttavat Hemo -kohtien b - SDHC: n ja SDHD.
Elektroninen virtaus sukkinaatista näiden kuljettajien läpi ja jopa lopullinen vastaanottaja, joka on happea, on kytketty 1: n synteesiin.5 ATP -molekyyliä jokaiselle elektroniselle vääntömomentille hengitysketjun fosforylaation kautta.
Inzima -viat
On raportoitu, että geenin mutaatiot, jotka koodaavat monimutkaisen sukkinaattidehydrogenaasin alayksikköä A.
Säätö
Kompleksisen sukkinaattidehydrogenaasin aktiivisuutta voidaan säädellä translaation jälkeisillä modifikaatioilla, kuten fosforylaatio ja asetylointi, Vaikka aktiivisen kohdan estäminen voi tapahtua myös.
Joidenkin lysiinijätteiden asetylointi voi vähentää tämän entsyymin aktiivisuutta ja tämä prosessi suoritetaan asetylaasin entsyymillä, joka tunnetaan nimellä SIRT3; Fosforylaatiolla on sama vaikutus entsyymiin.
Näiden modifikaatioiden lisäksi SDH -kompleksia säätelevät myös Krebs -syklin välittäjät, erityisesti Oksalastaatti ja sukunti. Oksalastaatti on voimakas estäjä, kun taas sukkinaatti suosii oksalasetaatin dissosiaatiota, joka toimii aktivaattorina.
Impulkinaatin puute -dehydrogenaasi
Sukkinaattivaje -dehydrogenaasi on poikkeavuus tai mitokondrioiden hengitysketjun häiriö. Tämän puutteen aiheuttavat SDHA (tai SDHAF1), SDHB-, SDHC- ja SDHD -mutaatiot.
Eri tutkimukset ovat osoittaneet homotsygoottisia ja heterotsygoottisia mutaatioita näissä geeneissä, erityisesti SDHA. Näiden geenien mutaatiot aiheuttavat aminohapposubstituutioita proteiinissa (missä.
Siksi aminohapposubstituutiot ja epänormaalin lyhyt proteiinikoodaus, aiheuttavat häiriöitä tai SDH -entsyymin muutoksia, jotka aiheuttavat vikaantumisen mitokondrioiden optimaalisessa kyvyssä energian tuottamiseksi. Tätä tutkijat kutsuvat mitokondrioiden hengitysketjun häiriöksi.
Tämä häiriö voidaan ilmaista ihmisen fenotyyppisesti monin tavoin. Tunnetuimpia ovat: kielellisen kehityksen puute tai puute, spastinen kvadrupleksi, lihasten tahattomat supistukset (dystonia), lihasheikkous ja sydänlihakset, muun muassa siihen liittyvät ongelmat.
Jotkut potilaat, joilla on sukkinaattivaje-dehydrogenaasi, voivat ilmentää Leightin tautia tai Kearns-Sairen oireyhtymää.
Kuinka dehydratoitunut sukkinaattivaje havaitaan?
Tietyt tutkimukset viittaavat laadullisten testien ja analyysin sekä kvantitatiivisen, entsymaattisen, hengitysketjun biokemiallisen analyysin käyttöä. Toisaalta toiset viittaavat tutkittavien alayksiköiden eksonien polymeraasiketjureaktion (PCR) täydelliseen laajentumiseen ja sitten vastaavaan sekvensointiin.
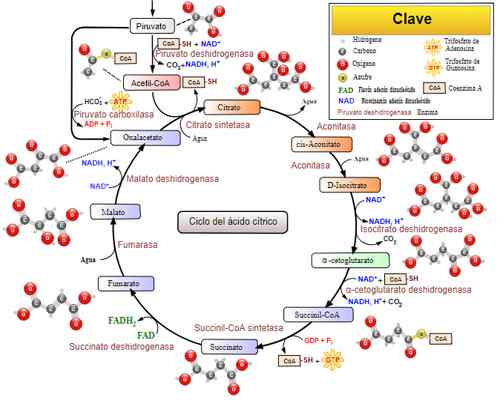 Trikarboksyylihapposykli (Krebs -sykli). Otettu ja muokattu julkaisusta: Narayanese, Wikiuserpedia, Yassinemrabet, Totobaggins (kääntänyt espanjaksi Alejandro Porto) [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)].
Trikarboksyylihapposykli (Krebs -sykli). Otettu ja muokattu julkaisusta: Narayanese, Wikiuserpedia, Yassinemrabet, Totobaggins (kääntänyt espanjaksi Alejandro Porto) [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)]. Liittyvät sairaudet
Mitokondrioiden hengitysketjun häiriöiden aiheuttamia fenotyyppisiä ekspressioita on sukkinaattivaje -dehydrogenaasin vuoksi. Kuitenkin, kun kyse on oireyhtymistä tai sairauksista, seuraavista puhutaan.
Voi palvella sinua: 8 tärkeintä biogeokemiallista sykliä (kuvaus)Leight -oireyhtymä
Se on progressiivinen neurologinen sairaus, joka liittyy mutaatioihin ydingenomissa (tässä tapauksessa dehydrogenaasin sukkinaatissa), jotka vaikuttavat pyruvaatin-dishytydrogenaasikompleksiin oksidatiiviseen fosforylaatioreitteeseen.
Oireet ilmestyvät ennen yksilön ensimmäistä vuotta, mutta epätyypillisissä tapauksissa ensimmäiset oireet murrosikäisen aikana on havaittu.
Yleisimmin havaittujen oireiden joukossa ovat: hypotonia, jolla on kefaalinen kontrolli, tahattomat liikkeet, toistuva oksentelu, hengitysvaikeudet, kyvyttömyys liikuttaa silmäsoluja, pyramidia ja ekstrapiramidaalisia merkkejä muun muassa muun muassa muun muassa muun muassa. Kohtaukset eivät ole kovin yleisiä.
On mahdollista, että tauti voidaan havaita synnytyksen diagnooseissa. Erityistä parannusta tai hoitoa ei tunneta, mutta jotkut asiantuntijat ehdottavat hoitoja tietyillä vitamiinilla tai kofaktoreilla.
Ruoansulatuskanavan stroma -kasvain (GIST)
Yleisesti nimeltään GIST, se on eräänlainen maha -suolikanavan kasvain, joka yleensä kehittyy sellaisilla alueilla kuin vatsa tai ohutsuoli. Uskotaan, että näiden syy johtuu tietystä erittäin erikoistuneista soluista, joita kutsutaan ICC -soluiksi tai Cajalin interstitiaalisoluiksi.
Muita näkökohtia GIST: n syytä ovat tietyntyyppisten geenien mutaatiot, jotka joidenkin kirjoittajien mukaan 90% kasvaimista. Mukana olevat geenit ovat: kit, pdgfra -geenit, dehydrogenaasi -sukkinaatti (SDH) - köyhä.
Suckerinaattidehydrogenaasi (SDH) - huono, esiintyy pääasiassa nuorilla naisilla, tuottaa mahalaukun kasvaimia ja suhteellisen taajuudella tuottaa metastaaseja imusolmukkeissa. Pieni prosenttiosuus tapahtuu lapsilla ja useimmissa tapauksissa se johtuu SDHB -alayksikön ilmaisun puutteesta.
Kearns-Sayre-oireyhtymä
On määritetty, että jotkut potilaat, joilla on sukkinaatti-dehydrogenaasin puutteita. Tämä sairaus liittyy mitokondriaalisiin häiriöihin, ja sille on ominaista silmäpallot liikkumisen puuttuminen.
Muita tämän taudin ominaisuuksia ovat pigmentti retinitis, kuuroisuus, kardiomyopatia ja keskushermosto -olosuhteet. Yleensä näitä oireita havaitaan ennen kuin potilas täyttää 20 vuotta vanhoja. Tämän tilan synnytystä diagnoosi ei tunneta.
Ei myöskään paranna tätä tautia. Hoito on palliatiivista, toisin sanoen se vain vähentää taudin vaikutuksia, ei parannuskeinoa. Toisaalta, vaikka se riippuu sairastuneiden elinten lukumäärästä ja saatuun lääketieteelliseen hoitoon, elinajanodote on suhteellisen normaali.
Viitteet
- Ackrell, b. -Lla., Kearney, E. B -., & Laulaja, t. P. (1978). [47] nisäkkäiden sukkinaattien dehydrogenaasi. Entsymologian menetelmissä (VOL. 53, s. 466-483). Akateeminen lehdistö.
- Brère, J. J -., Favier, J., Ghouzzi, V. JA., Djoudi, f., Benit, P., Gimenez, a. P., & Rustin, P. (2005). Sukkinaatti dehydrogenaasin vajaatoiminta ihmisessä. Solu- ja molekyylieläintieteet CMLS, 62 (19-20), 2317-2324.
- Cecchini, G., Schröder, minä., Gunalus, r. P., & Maklashina, ja. (2002). Escherichia colin sukkinaattidehydrogenaasi ja fumeratedeaktaasi. Biochimica et Biophysica Acta (BBA) -bioenergetics, 1553 (1-2), 140-157.
- Hatefi, ja., & Davis, K. -Lla. (1971). Sukkinaattidehydrogenaasi. Yllyttää. Puhdistus, molekyyliominaisuudet ja alarakenne. Biokemia, 10 (13), 2509-2516.
- Hederstedt, L. -Lla. R -. S., & Rutberg, L. -Lla. R -. S. (1981). Sukkinaatti dehydrogenaasi-A-vertaileva katsaus. Mikrobiologiset arvostelut, 45 (4), 542.
- Nelson, D. Lens., Lehninger, a. Lens., & Cox, M. M. (2008). Lehninger -biokemian periaatteet. Macmillan.
- Rutter, j., Winge, d. R -., & Schiffman, J. D -d. (2010). Sukkinaatti dehydrogenaasi-asemas, säätely ja rooli ihmisen diew: ssä. Mithochondrion, 10 (4), 393-401.
- « Välttämättömät rasvahappojen toiminnot, merkitys, nimikkeistö, esimerkit
- Sääntö T -ominaisuudet, niin että se palvelee esimerkkejä »