

Tilinlokki

- 1523
- 173
- Louis Moen
Tilinlaatikko Mitä ovat talkoideja?
Se Tilinlokki Ne ovat osastoja litteinä pussina, jotka sijaitsevat kloroplastien sisällä kasvisoluissa, syanobakteereissa ja leväissä. Ne on yleensä järjestetty rakenteeseen, jota kutsutaan grana -malleiksi Granum- Ja näyttää kasa kolikoita.
Tilakoideja pidetään kolmantena kloroplastien kalvojärjestelmänä, lukuun ottamatta mainitun organellen sisäistä ja ulkoista kalvoa. Tämän rakenteen kalvo erottuu kloroplasti -strooman talkoidin sisällä, ja siinä on sarja pigmenttejä ja proteiineja, jotka osallistuvat metabolisiin reitteihin.
Tilakoideissa on välttämättömiä biokemiallisia reaktioita fotosynteesille, prosessille, jolla kasvit vievät auringonvaloa ja muuttavat ne hiilihydraateiksi. Erityisesti heillä on tarvittavat koneet, jotka on ankkuroitu kalvoonsa auringonvalosta riippuvaisen vaiheen suorittamiseksi, joissa valo on loukussa ja muunnetaan energiaksi (ATP) ja NADPH: ksi.
Tilakoidien ominaisuudet
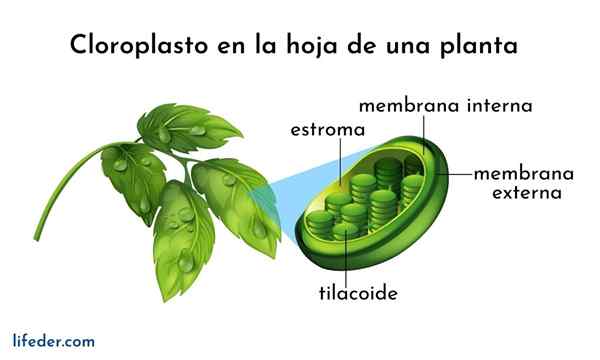
- Tilakoidit ovat sisäinen kolmen dimensioinen kalvokloroplastien kalvojärjestelmä. Täysin kypsät kloroplastit ovat 40 - 60 grana pinottu, halkaisija on välillä 0,3 - 0,6 um.
- Granaa muodostavien tilakoidien lukumäärä vaihtelee suuresti: alle 10 pussista kasveissa, jotka altistetaan riittävälle auringonvalolle, jopa 100 tilakoidiin kasveissa, jotka asuvat ympäristöissä, joilla on äärimmäinen varjo.
- Pinottu talkoidit on kytketty toisiinsa muodostaen jatkuvan osaston kloroplastien sisällä. Tilakoidin sisätilot on melko tilava vesipitoinen osasto.
- Tilakoidikalvo on välttämätön fotosynteesille, koska prosessin ensimmäinen vaihe tapahtuu siellä.
Tilinrakenne
Tilakoidit ovat rakenteita, jotka hallitsevat täysin kypsän kloroplastien sisällä. Jos perinteisessä optisessa mikroskooppissa esitetään kloroplasti, jyvälajeja voidaan havaita.
Nämä ovat talkoidipinoamista; Siksi näiden rakenteiden ensimmäisiä tarkkailijoita kutsuttiin "grana".
Elektronisen mikroskoopin avulla kuvaa voitiin laajentaa ja pääteltiin, että näiden jyvien luonne oli todella pinottu tilacoideja.
Tilakoidikalvon muodostuminen ja rakenne riippuu kloroplastien muodostumisesta, joka ei ole vielä eriytetty plastidium, joka tunnetaan nimellä Protoplastidio. Valon läsnäolo stimuloi muuntamista kloroplastiiksi ja myöhemmin pinottujen tilakoidien muodostuminen.
Voi palvella sinua: siittiöt: toiminnot, osat, elinkaariTilinsilakalvo
Kloroplasteissa ja syanobakteereissa Tilcoid -kalvo ei ole kosketuksessa sisäisen plasmamembraanin kanssa. Tilakoidikalvon muodostuminen alkaa kuitenkin sisäisen kalvon tunkeutumisella.
Syanobakteereissa ja tietyissä levälajeissa tilakoideja muodostetaan yhdellä lamellakerroksella. Sitä vastoin kypsissä kloroplasteissa löytyy monimutkaisempi järjestelmä.
Tässä viimeisessä ryhmässä voit erottaa kaksi olennaista osaa: Grana ja Stroma Lamella. Ensimmäinen koostuu pienistä pinotuista albumeista ja toinen vastaa näiden pinojen yhdistämisestä toisiinsa, rakenteen muodostaminen jatkuu: Tilcoidin luumen.
Kalvon lipidikoostumus
Kalvon muodostavat lipidit ovat erittäin erikoistuneita ja koostuvat lähes 80 %: sta diasyyliglyserolista galaktosyylistä: monogalaktosyylidiasyyliglyseroli ja diglactosyylidiasyyliglyseroli. Näillä galactolipideilla on erittäin tyydyttymättömät ketjut, jotka ovat tyypillisiä talkoideille.
Samoin tilakoidikalvo sisältää lipidejä, kuten fosfatidyyliglyserolia vähemmän suhteessa. Mainitut lipidit eivät ole jakautuneet homogeenisesti kalvon molemmissa kerroksissa; On tietty epäsymmetria, joka näyttää edistävän rakenteen toimintaa.
Membraaniproteiinikoostumus
Valosysteemit I ja II ovat hallitsevat proteiinikomponentit tässä kalvossa. Ne liittyvät sytokromi b -kompleksiin6F ja ATP -syntetaasi.
On havaittu, että suurin osa Photosystem II: n elementeistä sijaitsee pakattuissa Grana -kalvoissa, kun taas Photosystem I sijaitsee enimmäkseen antamattomissa talkoidikalvoissa. Eli molempien valosysteemien välillä on fyysinen erotus.
Nämä kompleksit sisältävät kattavat membraaniproteiinit, perifeeriset proteiinit, kofaktorit ja erilaiset pigmentit.
Tilinlahka
Tilakoidin sisustus koostuu vesipitoisesta ja paksusta aineesta, jonka koostumus on erilainen kuin strooma. Osallistu fotofosforylaatioon, protonien tallentaminen, jotka tuottavat protonemoottorin voiman ATP-synteesille. Tässä prosessissa luumen pH voi saavuttaa 4.
Malli -organismin lummen -proteoomassa Thalian Arabidopsis Yli 80 proteiinia on tunnistettu, mutta niiden toimintoja ei ole täysin selvitetty.
Lumen -proteiinit osallistuvat tilakoidisen biogeneesin säätelyyn sekä fotosynteettisten kompleksien, erityisesti fotosysteemin II ja NAD (P) H D Edrogensa, aktiivisuuteen ja korvaamiseen.
Se voi palvella sinua: Promielocito: Ominaisuudet, toiminnot ja patologiatTilakoidifunktiot
Vihannesten elintärkeä fotosynteesiprosessi alkaa Tilcoideissa. Kalvo, joka rajaa ne kloroplasti stromilla.
Fotosynteesin vaiheet
Fotosynteesi voidaan jakaa kahteen suureen vaiheeseen: vaaleat reaktiot ja tummat reaktiot.
Kuten nimestä voi päätellä. Huomaa, että ympäristö ei ole välttämätöntä olla "tumma", se on vain valosta riippumaton.
Ensimmäinen reaktioryhmä, "valo", esiintyy Tilcoidissa ja voidaan tiivistää seuraavasti: Light + klorofylli + 12 h2O + 12 NADP+ + 18 ADP + 18 pYllyttää à 6 o2 + 12 NADPH + 18 ATP.
Toinen reaktioryhmä esiintyy kloroplastien stroomassa ja ottaa ATP: n ja NADPH: n syntetisoidun ensimmäisen vaiheessa hiilidioksidihiilen vähentämiseksi glukoosiin (C6H12JOMPIKUMPI6-A. Toinen vaihe voidaan tiivistää: 12 NADPH + 18 ATP + 6 Co2 à c6H12JOMPIKUMPI6 + 12 NADP+ + 18 ADP + 18 pYllyttää + 6 h2JOMPIKUMPI.
Valosta riippuvainen vaihe
Kevyet reaktiot sisältävät sarjan rakenteita, jotka tunnetaan nimellä fotosysteemit, joita löytyy Tilcoid -kalvosta ja sisältävät 300 pigmenttimolekyylin sisällä, näiden klorofyllin joukossa.
Valosysteemiä on kahta tyyppiä: Ensimmäisellä valon imeytymishuipulla on 700 nanometriä ja tunnetaan nimellä P700, Kun taas toista kutsutaan p680. Molemmat on integroitu tilacoid -kalvoon.
Prosessi alkaa, kun yksi pigmenteistä imee fotonin ja tämä "pomppiminen" kohti muita pigmenttejä. Kun klorofyllimolekyyli absorboi valoa, elektroni hyppää ja toinen molekyyli absorboi sen. Elektronin menettänyt molekyyli on nyt hapettunut ja sillä on negatiivinen kuorma.
P680 Ota kevyttä energiaa klorofyllistä a. Tässä fotosysteemissä elektroni heitetään enemmän energiajärjestelmään ensisijaiseen elektronia vastaanottajaan.
Tämä elektroni putoaa fotosysteemiin I elektronikuljetinketjun kautta. Tämä hapettumis- ja pelkistysreaktiojärjestelmä on vastuussa protonien ja elektronien siirtämisestä molekyylistä toiseen.
Voi palvella sinua: lymfoblastitToisin sanoen, vedestä fotosystem II: n elektronien virtaus fotosysteemiin I ja NADPH.
Fotofosforylaatio
Osa tämän reaktiojärjestelmän tuottamista protoneista sijaitsee talkoidin sisällä (kutsutaan myös Tilcoid LUZ), joka luo kemiallisen gradientin, joka tuottaa protonemoottorin voiman.
Protonit liikkuvat tilakoidisesta tilasta stroomaan, suotuisasti sähkökemiallisen gradientin jälkeen; eli he jättävät talkoidin.
Protonien kulku ei kuitenkaan ole missään kalvossa, heidän on tehtävä se monimutkaisen entsymaattisen järjestelmän kautta, nimeltään ATP -synteesiesi.
Tämä protonin liike kohti Stromia. ATP -synteesiä valoa kutsutaan fotofosforylaatioksi.
Nämä mainitut vaiheet tapahtuvat samanaikaisesti: Photosystem II -klorofylli menettää elektronin ja sen on korvattava se elektronilla vesimolekyylin repeämästä; Photosystem I tarttuu valoon, hapettaa ja käynnistää elektronin, jonka NADP on kiinni+.
Photosystem I: n kadonnut elektroni korvataan Photosystem II: n tuloksella. Näitä yhdisteitä käytetään seuraavissa hiilen kiinnitysreaktioissa Calvin -syklissä.
Kehitys
Fotosynteesin kehitys happea -liitosprosessi salli elämän sellaisena kuin me sen tiedämme.
Ehdotetaan, että fotosynteesiä kehitettiin muutamia miljardeja vuosia sitten esi -isässä, joka johti nykyisiin syanobakteereihin, jotka perustuvat anksiseen fotosynteettiseen kompleksiin.
Ehdotetaan, että fotosynteesin kehitykseen liittyi kaksi välttämätöntä tapahtumaa: Photosystem P: n luominen680 ja sisäisen kalvojärjestelmän synty, ilman yhteyttä solukalvoon.
Tilacoides -muodostumiselle on välttämätön VIPP1 -proteiini. Itse asiassa tätä proteiinia on läsnä kasveissa, levässä ja syanobakteereissa, mutta puuttuvat bakteereissa, jotka suorittavat anoksisia fotosynteesiä.
Uskotaan, että tämä geeni voi johtua geenin kopioinnista syanobakteerien mahdollisessa esi -isässä. On vain yksi syanobakteerinen tapaus, joka kykenee suorittamaan happea fotosynteesiä ja jolla ei ole tilakoideja: laji Gloeobacter violaceus.