

Trombosytopoieesiprosessi, stimulantit, säätely
- 4186
- 805
- Arthur Terry II
Se trombosytopoieesi Se on verihiutaleiden koulutus- ja vapauttamisprosessi. Tämä prosessi suoritetaan luuytimessä sekä erytropoiees ja granulopoyesis. Verihiutaleiden muodostuminen käsittää kaksi vaihetta: megakariopoyesis ja trombosytopoieesi. Megakariopyeesi alkaa myeloidi -suvun edeltäjäsolusta kypsän megakariosyytin muodostumiseen.
Toisaalta trombosytopoieesi sisältää sarjan tapahtumia, joiden kautta megakariosyyttihäiriöt. Tämä solu vastaanottaa erilaisia signaaleja riippuen sivustosta, jossa se sijaitsee.
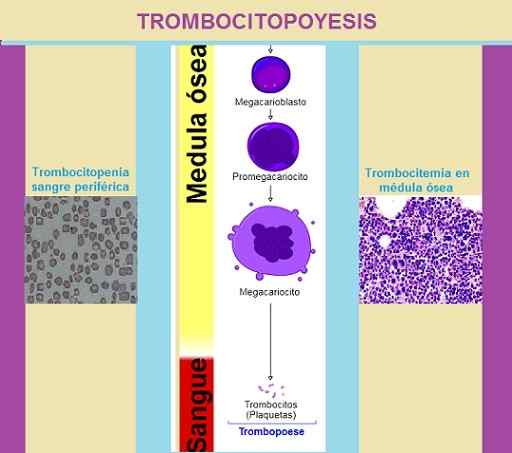
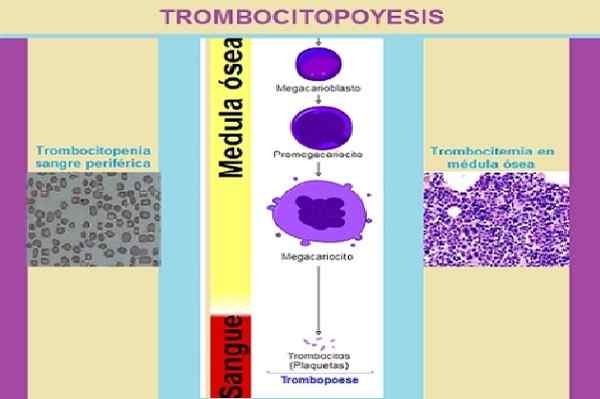 Trombosytopoyesifaasit. Kuvan suunnittelu: Marielsa Gil. Lähteet lukuista: a. Rad [cc by-sa 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)]/Ei koneeta luettavissa olevaa kirjailijaa. KGH oletetaan (tekijänoikeusvaatimusten perusteella). [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]/prof. Erhabor Osaro [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)]
Trombosytopoyesifaasit. Kuvan suunnittelu: Marielsa Gil. Lähteet lukuista: a. Rad [cc by-sa 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)]/Ei koneeta luettavissa olevaa kirjailijaa. KGH oletetaan (tekijänoikeusvaatimusten perusteella). [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]/prof. Erhabor Osaro [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Kun solu on osteoblastisen stromin sisällä.
Nämä aineet ovat von willebrand -tekijä, fibrinogeeni ja verisuonen endoteelin kasvutekijä. Kun aktivoidaan, proplaquettes -nimisen megakariosyytin sytoplasmiset pidentymiset pirstoutuvat aiheuttamaan proplaquetasia ja verihiutaleita.
Trombosytopoiees -sääntelyn sääntelyprosessin ansiosta homeostaasi on mahdollista ylläpitää verihiutaleiden kiertävän määrän suhteen. Trombosytopoyesin stimuloivina tekijöinä on trombopoietiinia, Interleuquina 3 (IL3), IL 6 ja IL 11. Ja koska estävät tekijät ovat verihiutaleiden tekijä 4 ja transformoiva kasvutekijä (TGF) β.
On olemassa useita sairauksia, joissa kiertävien verihiutaleiden lukumäärä muuttuu, samoin kuin niiden morfologia tai niiden toiminta. Nämä poikkeavuudet aiheuttavat vakavia ongelmia niistä, jotka kärsivät heistä, etenkin verenvuodot ja tromboosi, muun muassa.
[TOC]
Trombosytopoyesiprosessi
Verihiutaleiden muodostuminen voidaan jakaa kahteen prosessiin, ensimmäistä kutsutaan megakariosytopoyesiksi ja toiseksi trombosytopoyssiksi.
Kuten tiedetään, kaikki solulinjat tulevat pluripotentiaalisesta kantasolusta. Tämä solu eroaa kahden tyyppisissä progenitorisoluissa, yksi myeloidi -suvusta ja toinen imusolmukkeesta.
Myeloid-linjan progenitorisolusta 2-tyyppisiä soluja syntyy, megakariosyyttistä eritroidi-vanhempaa ja granulosyyttinen makrofagin progenitori.
MEGAKARICITIC-EREROID-PROGENItor-soluista, megakiaiosyyteistä ja punasoluista.
-Megakariosytopoyesis
Megakariosytopoyesi sisältää solujen erilaistumis- ja kypsymisprosessin Purskeinen yksikkö (BFU-MEG) Megakariosyytin muodostumiseen asti.
Cuf-gemm
Tämä solu syntyy kantasolusta ja siitä johdetaan granulosyyttisen makrofagin ja megaryosyyttisen jautroidisolujen progenitorisolut.
Bfu-meg
Tämä solu on megakariosyyttisen sarjan primitiivisin näyte. On suuri proliferatiivinen kapasiteetti. CD34+/HLADR -vastaanottimelle on ominaista esittämällä sen kalvossa-
CFU-MEG
Sen lisääntyvä kapasiteetti on pienempi kuin edellinen. Se on hiukan erilaistunut kuin edellinen ja sen kalvossa esitetään CD34+/HLADR -reseptori+
Promotecarioblast
Se mittaa 25 ja 50 um, se on epäsäännöllinen ja suuri ydin. Sytoplasma on hiukan basofiilistä ja sillä voi olla kevyt polychromasia. Voi olla 0 - 2 Nucleó.
Megakarioblast
Tälle solulle on ominaista pienempi koko kuin megakariosyytti (15-30 um), mutta paljon suurempi kuin muut solut. Siinä on yleensä bilobed näkyvä ydin, vaikka joskus se voi olla olemassa ilman lobulaatioita.
Kromatiini on rento ja useita nukleoleja arvostetaan. Sytoplasma on basofiilinen ja niukka.
Promisecariocito
Tälle solulle on ominaista polyiloboitu ja matala. Sytoplasma on runsaampi ja se erotetaan monikromaattisesti.
Voi palvella sinua: Heterotroph Nutrition: Ominaisuudet, vaiheet, tyypit, esimerkitMegakariosito
Tämä on suurin solu, sen mitat ovat välillä 40-60 µm, vaikka megakiaiosyytejä on nähty, että mitat ovat 100 um. Megakariosyytteillä on runsaasti sytoplasma, joka on yleensä eosinofiilinen. Sen ydin on suuri polyploidi ja siinä on useita lobulaatioita.
Tämän solun kypsymisprosessissa se on linjan ominaisuuksia, kuten spesifisten verihiutaleiden rakeiden (atsurofiilien) esiintyminen tai sytoskeleton tiettyjen komponenttien synteesi, kuten aktiini, tubuliini, filamiini, alfa-1-aktiiniini ja aktiiniini ja aktiini ja aktiini myosiini.
Heillä on myös solumembraanin tunkeutuminen, joka muodostaa kompleksisen kalvon rajajärjestelmän, joka ulottuu koko sytoplasmaan. Jälkimmäinen on erittäin tärkeä, koska se on verihiutaleiden kalvojen muodostumisen perusta.
Näiden solujen muut ominaisuudet ovat seuraavat:
- Spesifisten markkerien esiintyminen niiden kalvossa, kuten: glykoproteiini IIBIIIA, CD 41 ja CD 61 (fibrinogeenireseptorit), glykoproottinen kompleksi IB/V/IX, CD 42 (von Willebrand -tekijäreseptori).
- Endomitoosi: Prosessi, jossa solu kertoo DNA: nsa kaksinkertaiseksi jakamatta, prosessin kautta, jota kutsutaan abortiviksi myitoosiksi. Tämä prosessi toistetaan useissa sykleissä. Tämä antaa sille ominaisuuden olla suuri solu, joka tuottaa monia verihiutaleita.
- Sytoplasmisten pidentymisten esiintyminen, joka on samanlainen kuin pseudopodit.
Verihiutaleet
Ne ovat hyvin pieniä rakenteita, mittaa välillä 2-3 um, niillä ei ole ydintä ja niissä on 2 tyyppisiä rakeita, joita kutsutaan alfaksi ja tiheiksi. Kaikista mainituista soluista nämä ovat ainoat, jotka voidaan nähdä perifeerisessä verenkierrossa. Sen normaali arvo vaihtelee välillä 150.000 - 400.000 mm3. Hänen puoliintumisajansa on noin 8-11.
-Trombosytopoieesi
Jo kypsä megakariosyytti vastaa verihiutaleiden muodostamisesta ja vapauttamisesta. Megakariosyyttejä, jotka ovat lähellä verisuonten endoteeliä luuydin sinusoidissa.
ProPlaquettes -alueen uloin alue on pirstoutunut verihiutaleiden aiheuttamiseksi. Verihiutaleiden vapautumista tapahtuu verisuonissa, ja sitä auttaa verenkiertotorren voima. Tätä varten proplaquetan on ylitettävä endoteeliseinä.
Jotkut kirjoittajat sanovat, että proplaquetan ja verihiutaleiden välillä on välittäjävaihe, jotka ovat kutsuneet pre -peppereiksi. Tämä muutos proplaquetasta preplaquetaksi näyttää olevan palautuva prosessi.
Prepatches ovat suurempia kuin verihiutaleet ja niissä on diskoidinen muoto. Lopuksi niistä tulee verihiutaleita. Muutaman tunnin kuluttua megakariosyyttistä on syntynyt yhteensä 1000–5000 verihiutaleita.
Trombosytopoyesin stimulantit
Stimuloivien aineiden joukossa ovat kantasolun stimuloiva tekijä, Interleuquina 3, interleuquina 6, interleuquina 11 ja trombopoietiini.
Interleuquina 3
Tämä sytoquine puuttuu lisäämällä megakariosyyttisen suvun primitiivisimpien ja epäkypsien kantasolujen käyttöikää. Tämä tehdään estämällä näiden solujen apoptoosiprosessin tai ohjelmoidun solukuoleman.
Interleuquina 6
Se on tulehdusta edistävä interleuquina, joka esittelee viraston erilaisia tehtäviä. Yksi sen toiminnoista on stimuloida hematopoieettisten prekursorien synteesiä, joista on megakariosyyttisen linjan esiasteiden stimulaatio. Toimi CFU-Gemm: n erottelusta CFU-MEG: hen.
Se voi palvella sinua: Müeller Hinton Agar: Mikä on, perusta, valmistelu, käyttääInterlequina 11
Kuten trombopoietiini, se toimii koko megakariosytopoieesin prosessin tasolla, toisin sanoen pluripotentiaalisten solujen stimulaatiosta megakariosyyttien muodostumiseen.
Trombopoietiini
Tämä tärkeä hormoni syntetisoidaan pääasiassa maksassa ja toissijaisesti munuaisten ja luuytimen stromassa.
Trombopoietiini toimii luuytimessä, stimuloimalla megakariosyyttien ja verihiutaleiden muodostumista. Tämä sytoquine puuttuu megakariopoyesin ja trombosytopoysin kaikissa vaiheissa.
Uskotaan, että se myös stimuloi kaikkien solulinjojen kehitystä. Se myötävaikuttaa myös verihiutaleiden asianmukaiseen toimintaan.
Trombosytopoyesin säätely
Kuten mikä tahansa prosessi, trombosytopoieesia säädetään tietyillä ärsykkeillä. Jotkut suosivat verihiutaleiden muodostumista ja vapauttamista verenkiertoon ja toiset estävät prosessia. Nämä aineet syntetisoivat immuunijärjestelmän solut, luuytimen strooma ja endoteeliset retikulumisolut.
Säätelymekanismi aiheuttaa verihiutaleiden määrän pysyvän normaalilla tasolla verenkierrossa. Noin päivittäiset verihiutaleet ovat 10yksitoista.
Luuytimen strooman mikroympäristöllä on perustavanlaatuinen rooli trombosytopoyesin säätelyssä.
Kun megakariosyytti kypsyy, se liikkuu, siirtyen osastosta toiseen; Toisin sanoen se siirtyy osteoblastisesta osastosta verisuoneen seuraten kemotaktista gradienttia, nimeltään stroma -johdettu tekijä - 1.
Vaikka megakariosyytti on kosketuksessa osteoblastisen osaston (tyypin I kollageenin) komponenttien kanssa, proplaquettien muodostuminen estetään.
Tämä aktivoituu vasta, kun se on kosketuksissa Von Willebrand -tekijän ja fibrinogeenin kanssa, joka on läsnä verisuoniosaston solunulkoisessa matriisissa, kasvutekijöiden, kuten verisuonten endoteelin kasvutekijän (VEGF), vieressä (VEGF) vieressä.
-Trombopoietiini
Trombopoietiini eliminoidaan verihiutaleilla, kun se vangitaan MPL -vastaanottimen kautta.
Siksi, kun verihiutaleet lisäävät trombopoietiinia, vähenee korkean puhdistuksen vuoksi; Mutta kun verihiutaleet alentavat, sytoquinan plasma -arvo nousee ja stimuloi medullaa verihiutaleiden muodostumiseen ja vapautumiseen.
Luuytimessä syntetisoitua trombopoietiinia stimuloi veren verihiutaleiden lukumäärän lasku, mutta trombopoietiinin muodostumista maksaan stimuloidaan vasta, kun hepatosyytin Ashwell-Morell-reseptori aktivoidaan oletusvuorien läsnä ollessa.
Deialinisoidut verihiutaleet tulevat verihiutaleiden kärsimästä apoptoosiprosessista, kun ne ovat vangittuja ja poistetaan monosyytti-makrofagijärjestelmässä pernasuhteella.
-Estäjät
Verihiutaleiden harjoitusprosessin lopettavien aineiden sisällä verihiutaleiden tekijä 4 ja transformoiva kasvutekijä (TGF) β.
Verihiutaleiden tekijä 4
Tämä sytoquine sisältyy verihiutaleiden alfa -rakeisiin. SA tuntee myös fibroblastien kasvutekijä. Se vapautuu verihiutaleiden aggregaation aikana ja jarrujen megakariopoyesis.
Voi palvella sinua: Guanina: Ominaisuudet, rakenne, koulutus ja toiminnotTransformoiva kasvutekijä (TGF) β
Se syntetisoi erityyppiset solut, kuten makrofagit, dendriittisolut, verihiutaleet, fibroblastit, lymfosyytit, kondrosyytit ja astrosyytit, muun muassa. Sen toiminta liittyy erilaisten solujen erilaistumiseen, lisääntymiseen ja aktivointiin ja osallistuu myös megakariosytopoyjen estämiseen.
Trombosytopoyesin epätasapainon aiheuttamat sairaudet
On monia häiriöitä, jotka voivat muuttaa homeostaasia verihiutaleiden koulutuksen ja tuhoamisen suhteen. Jotkut heistä mainitaan alla.
Trombosytopenia amegakariosyyttinen synnynnäinen
Se on outo perinnöllinen patologia, jolle on ominaista mutaatio trombopoietiini/MPL -vastaanotinjärjestelmässä (TPO/MPL).
Siksi näillä potilailla megakariosyyttien ja verihiutaleiden muodostuminen on melkein nolla ja lopulta kehittyy medullaariseen aplasiaan, mikä osoittaa, että trombopoietiini on tärkeä kaikkien solulinjojen muodostumiselle.
Olennainen trombokytemia
Se on harvinainen patologia, jossa esiintyy trombosytopoieesien epätasapainoa, mikä aiheuttaa veressä jatkuvasti verihiutaleiden lukumäärän liioiteltua kasvua ja verihiutaleiden esiasteiden (megakariosyyttejä) hyperplastisen tuotannon luuytimessä luuytimessä.
Tämä tilanne voi aiheuttaa potilaan tromboosia tai verenvuotoja. Vika tapahtuu kantasolutasolla, joka on kallistettu kohti solulinjan liioiteltuja tuotantoa, tässä tapauksessa megakaryosyyttinen.
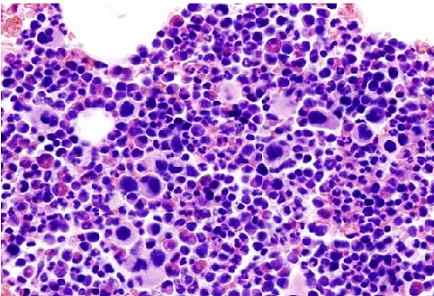 Olennainen trombokytemia (luuydinsälevy). Lähde: Ei konetta luettavissa olevaa kirjailijaa. KGH oletetaan (tekijänoikeusvaatimusten perusteella). [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]]
Olennainen trombokytemia (luuydinsälevy). Lähde: Ei konetta luettavissa olevaa kirjailijaa. KGH oletetaan (tekijänoikeusvaatimusten perusteella). [CC BY-SA 3.0 (http: // creativecommons.Org/lisenssit/by-SA/3.0/]] Trombosytopenia
Trombosytopeniaa kutsutaan verenvuorien vähentyneeksi lukumääränä veressä. Trombosytopenialla voi olla monia syitä, joista voidaan mainita: verihiutaleiden retentio pernassa, bakteeri -infektiot (JA. koli Enterohemorrgic) tai virusinfektiot (dengue, mononukleoosi).
Ne ilmestyvät myös autoimmuunisairauksiin, kuten systeeminen lupus erythematosus tai lääkitys alkuperät (sulfamidi, hepariini, antikonvulsiiviset hoidot).
Muita todennäköisiä syitä ovat verihiutaleiden tuotannon väheneminen tai verihiutaleiden tuhoamisen lisääntyminen.
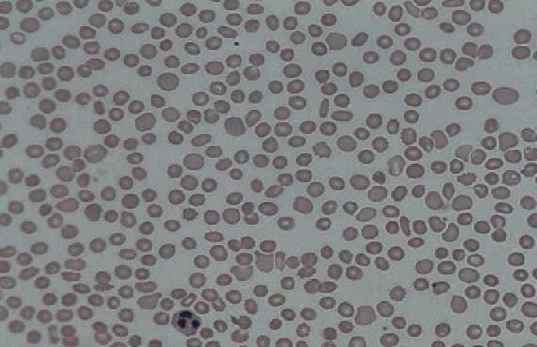 Perifeerinen verenkipu, jossa verihiutaleiden läsnäolo on pieni (trombosytopenia). Lähde: Prof. Erhabor Osaro [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)]
Perifeerinen verenkipu, jossa verihiutaleiden läsnäolo on pieni (trombosytopenia). Lähde: Prof. Erhabor Osaro [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Bernard-Soulier-oireyhtymä
Se on harvinainen perinnöllinen synnynnäinen sairaus. Sille on ominaista geneettisen muutoksen (mutaation) aiheuttama morfologia ja epänormaali toiminta, jossa VON Willebrand -tekijän vastaanotin (GPIB/IX) puuttuu.
Siksi hyytymisaikoja lisääntyy, trombosytopeniaa ja kiertävän makroplaquetin läsnäoloa.
Immunologinen trombosytopeeninen violetti
Tälle patologiselle tilalle on ominaista verihiutaleita vastaan tapahtuvien itsesianteiden muodostuminen, mikä aiheuttaa varhaisen tuhoamisen. Seurauksena on, että kiertävien verihiutaleiden lukumäärä vähenee huomattavasti.
Viitteet
- Heller P. Megakariosytopoyesi ja trombosytopoiees. Normaali hemostaasifysiologia. 2017; 21 (1): 7-9. Saatavana osoitteessa: sah.org.AR/aikakauslehti
- Mejía h, fuentes m. Immuunitrombosytopeeninen violetti. Rev Soc Bol Ped 2005; 44 (1): 64 - 8. Saatavana osoitteessa: Scielo.org.Bo/
- Bermejo e. Verihiutaleet. Normaali hemostaasifysiologia. 2017; 21 (1): 10-18. Saatavana osoitteessa: sah.org.AR
- Saavedra P, Vásquez G, González L. Interleucina-6: Ystävä tai vihollinen? Perusteet ymmärtää sen hyödyllisyyttä terapeuttisena tavoitteena. Iatreia, 2011; 24 (3): 157-166. Saatavana osoitteessa: Scielo.org.yhteistyö
- Ruiz-Gil W. Immuunitrombosytopeenisen purppuran diagnosointi ja hoito. Rev Med -perhed, 2015; 26 (4): 246-255. Saatavana osoitteessa: Scielo.org
- ”Trombopoyesis." Wikipedia, ilmainen tietosanakirja. 5. syyskuuta 2017, 20:02 UTC. 10. kesäkuuta 2019, 02:05 Saatavana osoitteessa: ES.Wikipedia.org
- Vidal J. Olennainen trombokytemia. Protokolla 16. Donostia -sairaala. 1-24. Saatavana osoitteessa: Osakidetza.Euskadi.Eus
- « ABO n yhteensopimattomuus-, interven- ja testijärjestelmä
- Oceanic Dorsals -ominaisuudet, miten ne muodostavat, esimerkkejä »