

Rasvahappojen beeta -hapettuminen, reaktiot, tuotteet, säätely
- 3438
- 1107
- Louis Moen
Se Rasvahappojen beeta -hapettuminen Se on rasvahappojen katabolismin (hajoamisen) reitti, jonka päätehtävänä näiden molekyylien yhteyksiin sisältyvän energian tuotanto tai "vapautuminen".
Tämä reitti löydettiin vuonna 1904 saksalaisen Franz Knoopin suorittamien kokeiden ansiosta, joka koostui rasvahappojen kokeellisista rotista, joiden lopullista metyyliryhmää oli modifioitu fenyyliryhmällä.
Rasvahappojen beetajärjestelmän hapettuminen (lähde: Arturo González Laguna [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta) Knoop odotti näiden "analogisten" rasvahappojen katabolismituotteita seuraamaan reittejä, jotka ovat samanlaisia kuin normaalien rasvahappojen hapettumisreitti (modifioimattomat alkuperäiskansojen). Hän havaitsi kuitenkin, että saatuissa tuotteissa oli eroja rasvahappojen hiiliatomien lukumäärästä riippuen.
Näiden tulosten avulla Knoop ehdotti, että hajoaminen tapahtui "askeleissa" alkaen "hyökkäys" β -hiilellä (sijainti 3 terminaalikarboksyyliryhmän suhteen) vapauttaen kahden hiiliatomin fragmentit.
Myöhemmin osoitettiin, että prosessi vaatii energiaa ATP: n muodossa, joka tapahtuu mitokondrioissa ja että kahden hiiliatomin fragmentit saapuvat Krebs-sykliin, kuten asetyyli-CoA.
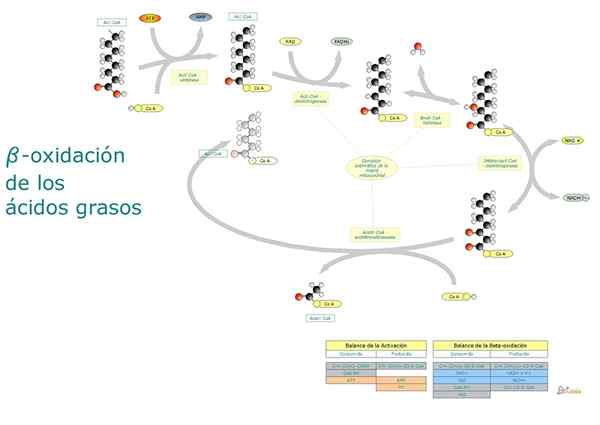
Lyhyesti sanottuna, rasvahappojen beeta -hapettuminen merkitsee terminaalikarboksyyliryhmän aktivointia, mitokondrioiden matriisin aktivoidun rasvahapon kuljetusta ja kahden ”porrastettua” hapettumista kahdessa karboksyyliryhmässä kahdessa hiilissä.
Kuten monet anaboliset ja kataboliset prosessit, tätä reittiä säännellään, koska se ansaitsee "varauksen" rasvahappojen mobilisoinnin, kun muut kataboliset reitit eivät riitä vastaamaan solu- ja kehon energian vaatimuksia.
[TOC]
Vaiheet ja reaktiot
Rasvahapot ovat pääosin sytosolissa, ne ovat jo biosynteettisiä reittejä tai rasvakerrostumista, jotka varastoidaan nautittuista elintarvikkeista (joiden on päästävä soluihin).
- Rasvahappojen aktivointi ja kuljetus mitokondrioihin
Rasvahappojen aktivointi vaatii ATP -molekyylin käytön, ja se liittyy aseita.
Se voi palvella sinua: biologian merkitysTätä aktivaatiota katalysoi entsyymien ryhmä, jota kutsutaan asetyyli-CoA-spesifinen pituus kunkin rasvan ketjun pituuden suhteen. Jotkut näistä entsyymeistä aktivoivat rasvahapot, kun ne kuljetetaan mitokondriaalimatriisiin, koska ne on upotettu ulkoiseen mitokondriaalikalvoon.
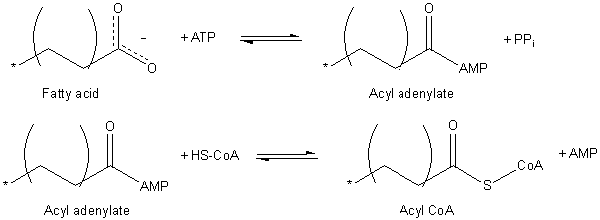 Rasvahappojen aktivointi (lähde: JAG123 englanniksi Wikipedia [julkinen alue] Wikimedia Commonsin kautta)
Rasvahappojen aktivointi (lähde: JAG123 englanniksi Wikipedia [julkinen alue] Wikimedia Commonsin kautta) Aktivointiprosessi tapahtuu kahdessa vaiheessa, ja se tuottaa ensin adenylaattiakyylin ATP -rasvahaposta, missä pyrofosfaattimolekyyli (PPI) vapautuu. ATP: n aktivoimaa karboksyyliryhmää hyökkää myöhemmin koentsyymin Tiol-ryhmäksi muodostamalla ACIL-CoA.
ACIL-CoA: n translokaatio mitokondriokalvon kautta saavutetaan karnitiinikuljetusnimisen kuljetusjärjestelmän ansiosta.
- Tyydyttyneiden rasvahappojen beeta -hapettuminen hiiliatomien parilla
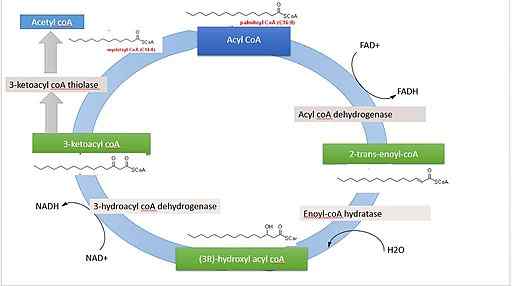
Rasvahappojen hajoaminen on syklinen reitti, koska kahden hiiliatomien kunkin fragmentin vapautumista seuraa välittömästi toinen, kunnes molekyylin kokonaispituus saavuttaa. Seuraavat reaktiot, jotka ovat osa tässä prosessissa, ovat seuraavat:
- Dehydraus.
- Kaksoissidoksen nesteytys.
- Hydroksyyliryhmän dehydraus.
- Pirstoutuminen asetyyli-CoA-molekyylin hyökkäykseen hiili-β: lla.
Reaktio 1: Ensimmäinen dehydraus
Se koostuu kaksoissidoksen muodostumisesta a -hiilen ja β -hiilen välillä poistamalla kaksi vetyatomia. Sitä katalysoi ACIL-CoA-dehydrogenaasientsyymi, joka muodostaa trans-transmolekyylin.
Reaktiot 2 ja 3: nesteytys ja dehydraus
Angel-Coatse katalysoi nesteytystä+.
Trans -valkoisen nesteytys.
Beetahapetuksen kolmessa ensimmäisessä reaktiossa tuotettu FADH2 ja NADH ovat uudelleenoksoituja.
Se voi palvella sinua: Ovogonias: Mitä ovat, kuvaus, morfologiaReaktio 4: pirstoutuminen
Jokainen hapettumisketosjakso, joka eliminoi kahden hiiliatomien päättymisen molekyylin.
Tätä reaktiota katalysoi β-kototoolaasi- tai tisolaasientsyymi, ja sen tuotteet ovat ACIL-CoA-molekyyli (rasvahappo, joka on aktivoitu kahdella vähemmän hiiliatomilla) ja yksi asetyyli-CoA: sta.
- Tyydyttyneiden rasvahappojen beeta -hapettuminen parittomalla määrällä hiiliatomeja
Näissä hiiliatomien parittoman määrän rasvahapoissa (jotka eivät ole kovin runsaasti) viimeisen hajoamisjakson molekyylissä on 5 hiiliatomia, joten sen pirstoutuminen tuottaa asetyyli-CoA-molekyylin (joka kulkee Krebsin sykliin) ja toisen Propionil-CoA.
Propionyyli-CoA: n on oltava karboksyloida (ATP ja bikarbonaattiriippuvainen reaktio) propionyyli-CoA-karboksylaasientsyymillä, joka muodostaa yhdisteen, joka tunnetaan nimellä D-metyylimalonyyli-CoA, joka on epimerisoitava sen muotoon "L".
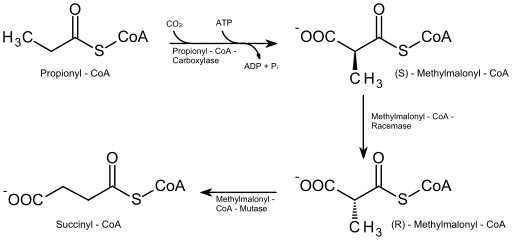 Parittomien lukujen rasvahappojen beetahapetus (lähde: Eleska [CC0] Wikimedia Commonsin kautta)
Parittomien lukujen rasvahappojen beetahapetus (lähde: Eleska [CC0] Wikimedia Commonsin kautta) Epimerisaatiosta johtuva yhdiste muunnetaan myöhemmin sucnyyli-CoA: ksi entsyymin L-metyylimalonyyli-CoA-mutaasin ja tämän molekyylin, samoin kuin asetyyli-CoA: n vaikutuksella, samoin kuin sitrushappojen sykli.
- Tyydyttymättömien rasvahappojen beeta -hapettuminen
Monissa solulipideissä on rasvahappiketjut tyydyttymättömyydellä, ts. Niillä on yksi tai useampi kaksoissidos hiiliatomien välillä.
Näiden rasvahappojen hapettuminen on hiukan erilainen kuin tyydyttyneiden rasvahappojen hapettuminen, koska kaksi ylimääräistä entsyymiä, isomeraasin vihaa ja 2,4-dieno-CoA-reduktaasia, vastaavat näiden tyydyttymättömyyden poistamisesta, jotta nämä rasvahapot voivat olla substraattia Entsyymi-CoA-hydratasa.
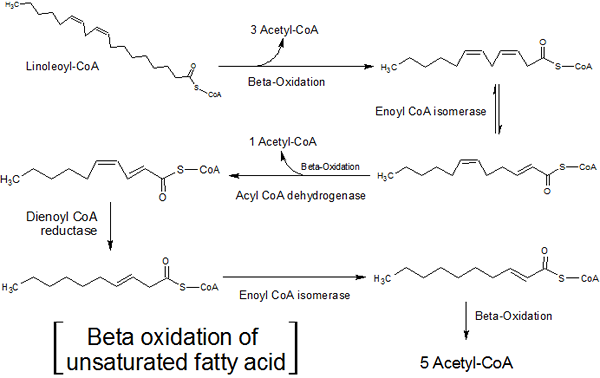 Tyydyttymättömien rasvahappojen beeta-hapettuminen (lähde: Hajime7Basketball [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Wikimedia Commonsin kautta)
Tyydyttymättömien rasvahappojen beeta-hapettuminen (lähde: Hajime7Basketball [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Wikimedia Commonsin kautta) Isomeraasihaput vaikuttaa monityydyttymättömiin rasvahappoihin (yhdellä tyydyttymättömyydellä), sillä välin entsyymi 2,4-dieno-CoA-reduktaasi reagoi monityydyttymättömien rasvahappojen kanssa (kahdella tai enemmän tyydyttymisellä).
- Beeta -ylimääräinen hapettuminen
Rasvahappojen beeta -hapettumista voi tapahtua myös muiden sytosolisten organelien, kuten peroksisomien sisällä.
Voi palvella sinua: Alstroemeria: Ominaisuudet, elinympäristö, hoito, lajitTämä reaktio tuottaa vetyperoksidia (happi on vähentynyt), yhdiste, joka eliminoi katlaasientsyymi, spesifinen näille organeleille.
Beetahapetustuotteet
Rasvahappojen hapettuminen tuottaa paljon enemmän energiaa kuin hiilihydraatin hajoaminen. Beetahapetuksen päätuote on asetyyli-CoA, joka on tuotettu reitin syklisen osan jokaisessa vaiheessa, muut tuotteet ovat kuitenkin:
- AMP, H+ ja Pyrofosfaatti (PPI), joka on tuotettu aktivoinnin aikana.
- Fadh2 ja nadh, jokaiselle tuotetulle asetyyli-CoA: lle.
- Succinil-coa, adp, pi, parittomille ketjuhapoille.
 Palmitiinihapon beetahapetus (lähde: 'Rojinbkht [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta)
Palmitiinihapon beetahapetus (lähde: 'Rojinbkht [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta) Jos tarkastellaan esimerkiksi palmitiinihapon (palmitaatin) täydellistä hapettumista, 16 hiiliatomien rasvahappoa, esiintyvän energian määrä vastaa enemmän tai vähemmän 129 ATP -molekyyliä, jotka tulevat 7 kierroksesta sen on suoritettava kiertää.
Säätö
Rasvahappojen beetahapetuksen säätely useimmissa soluissa riippuu energian saatavuudesta, paitsi hiilihydraatteihin, vaan samoihin rasvahappoihin.
Eläimet hallitsevat mobilisointia ja siksi rasvojen hajoamista hormonaalisilla ärsykkeillä, joita molekyylit, kuten esimerkiksi AMPC, säätelevät samanaikaisesti.
Maksassa, tärkeimmän rasvan hajoamisen elimen, malonyyli-CoA: n konsentraatio on erittäin tärkeä beetahapetuksen säätelyssä; Tämä on ensimmäinen rasvahappojen biosynteesireitille sitoutunut substraatti.
Kun malonyyli-CoA kertyy suuriin mittasuhteisiin, se edistää rasvahappojen biosynteesiä ja estää mitokondriaalista kuljetinta tai asyyli-karnitiinikuljetusta. Kun sen pitoisuus pienenee, estäminen lakkaa ja beetahapetus aktivoituu.
Viitteet
- Mathews, c., Van Holde, K., & Ahern, k. (2000). Biokemia (3. ed.-A. San Francisco, Kalifornia: Pearson.
- Nelson, D. Lens., & Cox, M. M. (2009). Lehninger -biokemian periaatteet. Omega -versiot (5. ed.-A.
- Rawn, j. D -d. (1998). Biokemia. Burlington, Massachusetts: Neil Patterson Publishers.
- Schulz, H. (1991). Rasvahappojen beeta -hapettuminen. Biochimica et Biophysica Acta, 1081, 109-120.
- Schulz, H. (1994). Rasvahappojen hapettumisen säätely sydämessä. Kriittinen arviointi, 165-171.
- Schulz, H., & Kunau, W. (1987). Tyydyttymättömien rasvahappojen beeta-hapettuminen: Revice-reitti. Piikit, 403-406.