

Glioksylaatit syklin ominaisuudet, reaktiot, säätely, toiminnot
- 4179
- 222
- Sheldon Kuhn
Hän Glioksylaattisykli Se on aineenvaihdunnan reitti, joka on läsnä kasveissa, joissain mikro -organismeissa ja selkärangattomien eläimissä (puuttuvat kaikissa selkärankaisissa), joiden kautta nämä organismit voivat muuttaa rasvat hiilihydraatteiksi (sokereiksi).
Tämä reitti löydettiin vuonna 1957, kun taas Kornberg, Krebs ja Beevers yrittivät selvittää kuinka bakteerit kuten Escherichia coli Ne voivat kasvaa asetaatin läsnä ollessa ainoana hiilen lähteenä ja kuinka taimet Tártagon itävyydessä (Ricinus communis) He voivat muuttaa rasvat hiilihydraateiksi.
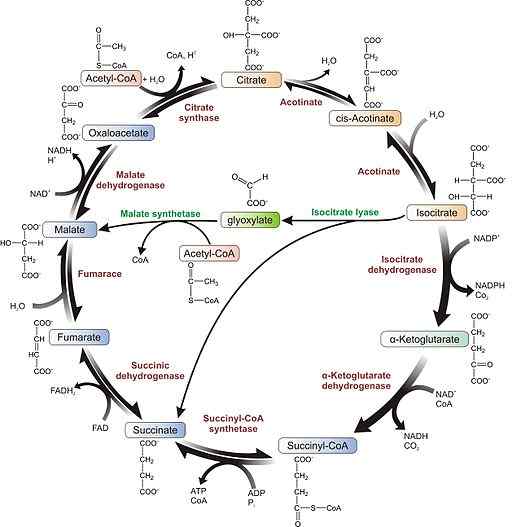
 Glioksilaattisyklijärjestelmä (lähde: Agrotman [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Wikimedia Commonsin kautta)
Glioksilaattisyklijärjestelmä (lähde: Agrotman [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Wikimedia Commonsin kautta) Näiden kolmen tutkijan tutkimukset johtivat kahteen Liasa-isositraatin ja pahan syntaasin nimeltään entsyymiin, jotka yhdessä Krebs-syklin entsyymien kanssa sallivat sukkinaattisynteesin kahdesta asetyyli-CoA-molekyylistä.
Näin tuotettu sukkinaatti muunnetaan Malatoksi trikarboksyylihapposyklin kautta, ja sitä voidaan myöhemmin käyttää glukoosin tuotantoon gluconeogeneesillä.
Tämä reitti tapahtuu kasveissa erityisissä organeleissa, joita kutsutaan glioksisomiksi, ja se on välttämätöntä taimien selviytymiselle itämisen varhaisessa vaiheessa.
[TOC]
Ominaisuudet
Glioksylaattireittiä voidaan harkita KREBS -syklin "modifikaationa", erolla, että ensimmäisessä ei tapahdu oksidatiivista dekarboksylaatiota, vaan että neljän hiiliatomien dikarboksyylihappot voidaan muodostaa kahdesta asetaattiyksiköstä kahdesta atomista kahdesta hiilistä.
Tätä glioksylaattisyklin ominaispiirteitä on kuvattu muotona, jonka joidenkin organismien on vältettävä (ohitus ") hiiliatomien menetystä hiilidioksidin muodossa, joka tunnistaa Krebs -syklin.
Kasveissa glioksylaattisykli tapahtuu joidenkin sytosolisten organelien sisällä, jota ympäröivät yksinkertainen kalvo, joka tunnetaan nimellä glyxisomit. Muissa organismeissa, kuten hiivat ja levät, toisaalta tämä reitti tapahtuu sytosolissa.
Glioksisoomat ovat rakenteellisesti samanlaisia kuin peroksisomit (jotkut kirjoittajat pitävät niitä ”erikoistuneiden peroksisomien”), toiset organelit, jotka vastaavat rasvahappojen β-hapettumista ja reaktiivisten happilajien eliminointia eukaryoottisissa organismeissa.
Se voi palvella sinua: gangliasidosSisällä rasvahapot hapettuu asetyyli-CoA: n tuottamiseksi, joka tiivistetään myöhemmin neljän hiiliatomin yhdisteissä. Nämä yhdisteet kuljetetaan selektiivisesti mitokondrioihin, joissa ne muunnetaan Malatoksi tai kuljetetaan sytosoliin päästäkseen glukoneogeeniseen reittiin (glukoosin synteesi).
Entsyymit, jotka on jaettu glioksylaattireitin ja trikarboksyylihapposyklin välillä, ovat mitokondrioissa ja glioksisomissa isoentsyymeinä, mikä tarkoittaa, että molemmat reitit toimivat enemmän tai vähemmän itsenäisesti itsenäisesti yksi muista.
Glioksisomien esiintyminen
Glioksisoomia ei ole pysyvästi kasvikudoksissa. Niitä on erityisen runsaasti oleagiinisten siementen itävyyden aikana, joilla on vähän fotosynteettistä kykyä tuottaa tarvitsemansa hiilihydraatit.
Täysin kehittyneissä kasveissa heidän osallistuminen rasvan aineenvaihduntaan ei ole niin välttämätöntä, koska sokerit saadaan pääasiassa fotosynteesillä.
Reaktiot
Asetaatti rasvahappojen hajoamisesta toimii rikkaana energian polttoaineena ja fosfoenolypyruvaatin lähteenä glukoosisynteesille glukonogeneesin avulla. Prosessi tapahtuu seuraavasti:
Glioksilaattisyklivaihit
1- Glioksylaattireitti, samanlainen kuin Krebs-sykli, alkaa asetyyli-CoA-molekyylin kondensaatiolla toisen oksalasetaatin kanssa sitraatin suorittamiseksi, entsyymin sitraattisyntaasin katalysoimaa reaktiota.
2- aconitosa-entsyymi muuntaa tämän sitraatin isositraatiksi.
3- Isocitratoa käytetään Liasa-isositraattientsyymin suklinaaliyhdisteiden ja glioksylaatin substraattina.
 Liasa -isositraattientsyymin molekyylirakenne (lähde: VrabiochemHW [CC0] Wikimedia Commonsin kautta)
Liasa -isositraattientsyymin molekyylirakenne (lähde: VrabiochemHW [CC0] Wikimedia Commonsin kautta) 4- Malato-syntasa-entsyymi ottaa glioksylaatin tuottaa pahaa kondensaation kautta toisen asetyyli-CoA-molekyylin avulla.
Voi palvella sinua: Orgaaniset biomolekyylit: ominaisuudet, toiminnot ja esimerkit5- paha dehydrogenaasi muunnetaan pahan oksalasaatiksi ja sanottu yhdiste voi toimia edeltäjänä glukoneogeeniselle reitille tai tiivistää toisen asetyyli-CoA: n kanssa syklin uudelleen käynnistämiseksi jälleen kerran uudelleen.
6- Tuotettu sukkinaali voidaan myös muuntaa fumaraatiksi ja tämä Malatoksi, tarjoamalla enemmän oksalasetaattimolekyylejä glukoosin muodostumista varten. Muutoin tämä molekyyli voidaan viedä myös mitokondrioihin työskentelemään Krebs -syklissä.
Oksalastaatti tulee glukoneogeeniseen reittiin glukoosituotantoon sen muuntamisen ansiosta fosfoenoliruvaatiksi, jota katalysoi entsyymi fosfoenolpiruvaatti karboksiquinaasi.
Säätö
Koska glioksylaatti- ja trikarboksyylihappojen syklillä on lukuisia välittäjiä keskenään, näiden kahden välillä on koordinoitu säätely.
Lisäksi on välttämätöntä, että kontrollimekanismeja on, koska glukoosin ja muiden heksaosien synteesi asetyyli-CoA: sta (rasvan hajoamisesta) merkitsee ainakin neljän reitin osallistumista:
- Rasvahappojen β-hapettuminen, joka tuottaa asetyyli-CoA-molekyylit, jotka ovat välttämättömiä sekä Krebs-syklille ja että kasveissa se tapahtuu glioksisomeissa.
- Glioksylaattisykli, joka esiintyy myös glioksisomeissa ja kuten todettiin, tuottaa välittäjiä, kuten sukkinaattia, pahaa ja oksalastaattia.
- Krebs -sykli, joka tapahtuu mitokondrioissa ja jossa välittäjät sukkina, paha ja oksalasaatti tapahtuu myös.
- Glukoneogeneesi, joka esiintyy sytosolissa ja pohtii oksalasetaatin käyttöä, muuttui fosfoenolypyruvaatiksi glukoosin syntetisoimiseksi.
Tärkein kontrollikohta on entsyymissä isositraattidehydrogenaasi, jonka säätely merkitsee kovalenttisen modifikaation lisäämällä tai poistamalla fosfaattiryhmä.
Kun entsyymi fosforyloituu, se on passiivinen, joten isosyytraatti on suunnattu kohti glukoosin tuotantoreittiä.
Funktiot
Kasvien osalta glioksylaattisykli on olennainen, etenkin itämisprosessin aikana, koska siemeniin varastoitujen rasvojen hajoamista käytetään glukoosisynteesiin huonosti kehitetyissä fotosyntegisesti puhuvissa kudoksissa.
Voi palvella sinua: glykogeeni: rakenne, synteesi, hajoaminen, toiminnotGlukoosia käytetään energian hankkimislähteenä ATP: n muodossa tai monimutkaisempien hiilihydraattien muodostumiseen rakennefunktioilla, mutta jotkut glioksylaattireitin aikana syntyneistä välittäjistä voivat myös palvella muiden solukomponenttien synteesitarkoituksia.
Mikro -organismeissa
Mikro -organismien glioksylaattisyklin päätehtävä on tarjota "vaihtoehtoinen" metabolinen reitti, joten mikro -organismit pystyvät hyödyntämään muita hiili- ja energialähteitä niiden kasvulle.
Näin on bakteerien tapaus Escherichia coli, jossa joidenkin glykolyysin ja sytoon happosyklin vähenemisen joidenkin välittäjien tasot (isositraatti, 3-fosfoglyserpaatti, pyruvaatti, fosfoenolypyruvaatti ja oksalasetaatti), entsyymi-isositraatti-dehydrogenaasi (joka osallistuu Krebs-sykliin) inhibidoitua ja estämää ja estämää on, suunnattu kohti glioksylaatin reittiä.
Jos tämä reitti on aktiivinen silloin, kun bakteerit kasvavat esimerkiksi väliaineessa, joka on runsaasti asetaattia, tätä metaboliittia voidaan käyttää syntetisoimaan neljän hiiliatomin karboksyylihappoja, jotka myöhemmin voivat johtaa energian hiilihydraattien muodostumiseen.
Esimerkiksi muille organismeille, kuten sienille.
Viitteet
- Dey, P., & Harborne, J. (1977). Kasvien biokemia. San Diego, Kalifornia: Academic Press.
- S. S. -Lla. (2006). Glyoksylaattisyklin tarkistaminen: Vaihtoehtoiset reitit mikrobiasetaatin assimilaatioon. Mikrobiologia Molecular, 61 (2), 274-276.
- Garrett, r., & Grisham, c. (2010). Biokemia (4. ed.-A. Boston, USA: Brooks/Cole. Cengage -oppiminen.
- Lorenz, m. C., & Fink, g. R -. (2001). Sienen virulenssiin tarvitaan glyksylaattisykli. Luonto, 412, 83-86.
- Mathews, c., Van Holde, K., & Ahern, k. (2000). Biokemia (3. ed.-A. San Francisco, Kalifornia: Pearson.
- Rawn, j. D -d. (1998). Biokemia. Burlington, Massachusetts: Neil Patterson Publishers.
- Vallarino, J. G., & Osorio, S. (2019). Orgaaniset hapot. Hedelmien ja vihannesten jälkeisessä fysiologiassa ja biokemiassa (PP. 207-224). Elsevier Inc.
- « Mesohyppus -ominaisuudet, lajit, lisääntyminen, ravitsemus
- Paquimé -kulttuurin alkuperä, sijainti, ominaispiirteet, uskonto »