

Arhaea -alue

- 5099
- 1029
- Shawn Stanton II
 Arkaat -kuva
Arkaat -kuva Mikä on Archaea -verkkotunnus?
Hän Arhaea -alue o Archaea Kingdom edustaa yhtä kolmesta elämän alueesta. Se koostuu hyvin samanlaisista mikroskooppisista yksisoluisista organismeista ja samalla hyvin erilainen kuin bakteerit ja eukaryootit monilla näkökohdilla.
Tämän ryhmän olemassaolo osoitettiin äskettäin, enemmän tai vähemmän vuoden 1970 lopulla, nimeltään myös arkeobakteerit.
WOEE- ja yhteistyökumppaneiden tutkimukset perustuivat vertaileviin fylogeneettisiin analyyseihin kaikkien elävien olentojen ribosomaalisten RNA: ien sekvenssien välillä, ja kolmen domeenin käsite ylläpidetään huolimatta valtavista sekvensseistä, joita edelleen lisätään ribosomaalisten RNA (tietokantojen) luetteloihin (tietokannat).
Nämä tutkimukset onnistuivat vilaamaan mahdollisuutta, että kaarit ovat eukaryootien veljesyhmä huolimatta niiden yhtäläisyyksistä prokaryootiensa (bakteerien) kanssa, jotta ne voisivat edustaa "menetettyä yhteyttä" prokaryootien ja eukaryootien välillä.
Uskotaan, että nämä mikro -organismit edustavat yli 20% kaikista valtameren vesien olemassa olevista prokaryooteista, välillä 1 - 5% maaperässä ja hallitsevin ryhmä meren sedimenteissä ja geotermisissä elinympäristöissä.
Kaarille on ominaista asumista ”äärimmäisissä” olosuhteissa, kuten kuumissa lähteissä, suolaissa, ympäristöissä, joissa on erittäin korkeat lämpötilat ja erittäin hapan pH, epämiellyttävä kohtaa, joissa happipitoisuus on hyvin pienentynyt tai nolla, jne.
Evoluutioperä ja rFylogeneettiset aliesit
Woesen ja yhteistyökumppaneiden ehdottamien elämän kolmen elämän alueen mahdollisesta alkuperästä on kolme teoriaa:
- Bakteerit erottuivat ensin, muodostaen suvun, joka tuotti kaaria ja eukaryooteja
- ”Proto-eucariotic” -linja poikkesi täysin proboottisesta suvusta (bakteerien ja kaarien linja)
- Kaarit erottuivat suvusta, joka myöhemmin johti eukaryooteihin ja bakteereihin
Vuonna 1989 kaksi tutkijaa, Gogarten ja Iwabe, ehdottivat itsenäisesti tapaa fylogeneettisesti analysoida kaikkia eläviä organismeja (mahdotonta tehdä ainutlaatuisten geenisekvenssien tutkimuksista).
IWABE käytti geenien kopioinnin "varhaisissa" tapahtumien geenien sekvenssien analysointia, "juurruttaen" elämänpuun pidennystekijöiden papukaijageenien sekvenssin vertailusta.
Pidentymiskertoimet ovat GTP: tä sitovia proteiineja, jotka osallistuvat translaatioon, erityisesti siirron RNA -molekyylien liitossa, joka on amioasiloitu ribosomeihin ja peptidiilien siirron siirtymisessä.
Kolmen ryhmän sekvenssien vertailujen tulosten mukaan kaarien pidentymistekijöiden koodien geenit ovat samankaltaisempia kuin eukaryoottisten organismien kuin bakteerien tulokset.
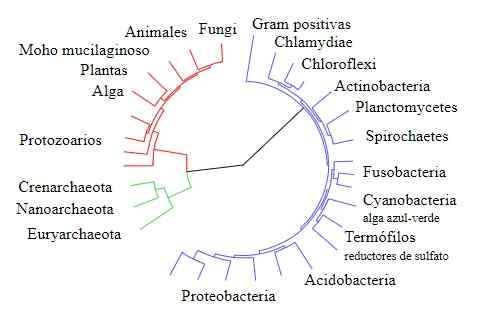 Elämän puu
Elämän puu Toisaalta Gogarten vertasi muiden päällekkäisyystapahtumien tuottamien geenien sekvenssejä, erityisesti sellaisia, jotka koodaavat tyypin V tyypin ja tyypin F alayksiköt, joita löytyy kaareista/eukaryooteista ja bakteereissa, vastaavasti, vastaavasti bakteereissa, vastaavasti bakteereissa, vastaavasti bakteereissa.
Gogartenin saamat tulokset, kuten edellä ilmoitetut, osoittavat, että nämä kaarien geenit (jotka väitetään syntyneet geneettisten päällekkäisyyksien tapahtumista) liittyvät läheisemmin eukaryooteihin kuin niiden bakteerien kanssa.
Näitä argumentteja tuki myös muiden tutkijoiden suorittama analyysi useita vuosia myöhemmin, jotka käyttivät sekvenssejä toisesta kaksoiskappaleesta, aminokasilisynteesasinsiirrosta, vahvistaen käsitystä kaarien ja eukaryootien välisestä läheisyydestä ".
Se voi palvella sinua: Selenito -liemi: Mikä on, perusta, valmistelu, käyttöWoese Tree
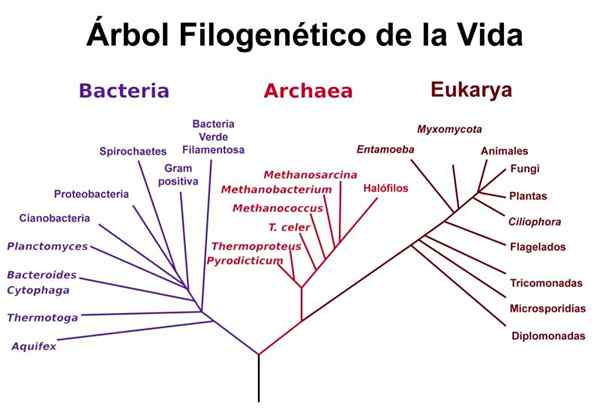 Woesen elämäpuu
Woesen elämäpuu Woese käytti Gogartenin ja Iwaben ja muiden ribosomaalisten RNA -sekvensseihin liittyvien tutkimusten suorittamia analyysejä ehdottaakseen sen elämänpuun "versiota", missä on selvää, että kaarit ja eukaryootit muodostavat "veljekset", jotka on erotettu bakteereista, jopa bakteereista Vaikka kaarien ja bakteerien väliset ribosomaaliset RNA -sekvenssit ovat samankaltaisempia kuin toiset.
Archaea -verkkotunnusominaisuudet
Kaaret tunnetaan joihinkin erityisistä (omista) ominaisuuksista ja myös ominaisuuksien "yhdistelmien" esittämisestä, joiden ajateltiin aikoinaan olevan yksinoikeudella bakteereille tai eukaryoottisille organismeille.
- Kuten bakteerit, kaarit Ne ovat prokaryoottisia virastoja, Toisin sanoen geneettisen materiaalin sisällä ei ole lukittu kalvolla (niillä ei ole ydintä), eikä sytoplasmisia kalvojen organeleja ole.
- Ne ovat yleensä bakteerien samanlaisia mikro -organismeja, niiden DNA on a Pyöreä kromosomi ja joitain pienempiä pyöreitä fragmentteja, jotka tunnetaan nimellä Plasmidit.
- He jakavat bakteerien kanssa Topoisomeraasi- ja retkien entsyymit Samanlainen, joka edustaa "epäsuoraa" todistetta "läheisyydestä" molempien organismiryhmien kromosomaalisen rakenteen suhteen.
- Archaeas -geenit osoittavat kuitenkin hienoja Homologia monien eukaryootien kanssa, etenkin niiden kanssa, joiden löytö on peräisin antibioottien tutkimuksista.
- Kone replikointi, transkriptio ja käännös eukaryootit ja kaarit ovat hyvin samanlaisia, etenkin entsyymin ADN -polymeraasin suhteen.
- Sen koodaavat geenit proteiinille Heillä ei ole introneja (Mutta muut tekevät), toisin kuin eukaryoottiset geenit. Lisäksi kaareissa on proteiineja, jotka ovat samanlaisia kuin niiden DNA: hon liittyvät histonit, läsnä eukaryooteissa ja puuttuvat bakteereissa.
- Niille on ominaista läsnäolo Isoprenilin eetterillipidit Solukalvoissa, samoin kuin asyyli-lipidien ja rasvahapposynteesasin puuttuessa.
- Yksi sen RNA -polymeraasientsyymin alayksiköistä on jaettu ja sen lähettiläs -RNA: lla, samoin kuin bakteereissa, ei ole "kapuchasia" (englannista lupa) Sen päissä 5 '.
- Heillä on hyvin erityinen antibioottiherkkyys ja heillä on Tyypin II rajoitusentsyymit hyvin samanlainen kuin bakteereille kuvattu.
- Toinen tärkeä ominaisuus liittyy siihen tosiasiaan, että suurella osalla kaareista on soluseinä, Mutta toisin kuin bakteerit, tämä ei koostu peptidoglykaanista.
Sen kalvolipidien ominaisuudet
Archaean -kalvolipidit eroavat huomattavasti bakteereissa ja eukaryoottisissa organismeissa, ja tätä on pidetty erittäin tärkeänä differentiaaliominaisuutena.
Suurin ero näiden amfipaattisten molekyylien (hydrofiilisen polaarisen pään ja hydrofobisen apolarin kanssa) välillä on, että glyserolin osan ja rasvahappojen ketjujen välinen liitto kaarien lipideissä on eetterilinkin kautta, kun taas bakteereissa ja eukaryoteissa vastaa esterilinkkiä.
Toinen tärkeä ero on, että kaareissa on lipidejä, joissa on rasvahappoja, joille on ominaista erittäin haarautuneiden isopreniliketjujen läsnäolo metyyliryhmien kanssa, sillä välin eukaryooteissa ja bakteereissa on pääasiassa rasvahappoja ei -haarautuneista ketjuista.
Eukaryootien ja bakteerien lipidit "rakennetaan" glyserolirunkoon, johon rasvahappiketjut esteröidaan hiiliatomeja 1 ja 2 vastaavissa asennoissa, mutta kaarissa glyserolieetterit sisältävät happoja, jotka sisältävät happoja rasvapaikoissa 2 ja 3.
Voi palvella sinua: Río Negro -pohja ja eläimistö: edustavat lajitToinen ero kalvolipidien suhteen liittyy sen biosynteettiseen reittiin, koska jotkut entsyymit ovat myös erilaisia kaareissa.
Esimerkiksi joillakin kaarilajeilla on bifunktionaalinen siirto prenil-entsyymi, joka vastaa esiasteiden tarjoamisesta sekä escualeenin synteesille että glyserilillipidi-isaprensoidien synteesille. Bakteereissa ja eukaryooteissa nämä toiminnot suoritetaan erillisillä entsyymeillä.
Kaarien luokittelu
Kaarien ribosomalien pienten alayksiköiden sekvenssien tietojen mukaan tämä ryhmä on jaettu pääasiassa kahteen ”reunaan”, jotka tunnetaan nimellä reuna Crenarchaeota ja reuna Euryarchaeta, jonka jäsenet ovat ennen kaikkea viljeltyjä kaaria In vitro.
Monia äskettäin kuvattuja kaaria ei kuitenkaan ole viljelty In vitro Ja ne liittyvät vain kaukaisesti sekvensseihin, jotka on eristetty laboratorioissa pidetyistä lajeista.
Reuna Crenarchaeota
 Termofiiliset bakteerit, crenarchaeota -reuna
Termofiiliset bakteerit, crenarchaeota -reuna Tämä ryhmä koostuu pääasiassa hypertermofiili- ja termasidofiilisten kaareiden lajeista, toisin sanoen ne kaareiden sukupuolet, jotka asuvat hävittämättömät ympäristöt, joissa on lämpöolosuhteet ja äärimmäinen pH.
Se koostuu yhdestä taksonomisesta luokasta, joka tunnetaan nimellä Teemooprotei, joiden sisällä ovat seuraavat viisi taksonomista tilausta: Acidilobales, Desulfurokokki, Fervicoccales, Sulfolobales ja Termoproteaalit.
Esimerkki joistakin näihin luokkiin kuuluvista genreistä voi olla genrejä Sulfolobus, Desulforococcus, Pyrodikrium, Teemo ja The thefilum.
Reuna Euryarchaeta
 Metanosarcina barkeri fusaro, euryarchaota filo
Metanosarcina barkeri fusaro, euryarchaota filo Tämän ryhmän jäsenillä on hiukan laajempi ekologinen alue, joka pystyy löytämään jonkin verran hypertermofiilisiä, metenogeenisiä, halofiili- ja jopa termofiilisiä mittareita, nimidikaareja, rikkiviikoiinta, rautaoksidaateja ja joitain organrofiilejä ja joitain organotrofiilejä.
Euriarqueotasin kuvattu taksonomiset luokat ovat kahdeksan ja tunnetaan nimellä Metania, Metanokokki, Metanobakteerit, Metanomikrobia, Archaeglobi, Halpa, Termokokit ja TEMOPLASMATA.
Monet tähän ryhmään kuuluvista kaareista jakautuvat laajasti, koska ne ovat maaperässä, sedimenteissä ja merivesissä sekä kuvattuissa äärimmäisissä ympäristöissä.
Reuna Thaumarhaeota
 Axinella -polypoidit, thaumarhaeota filo. Lähde: Liné1, CC BY-SA 3.0, Wikimedia Commons
Axinella -polypoidit, thaumarhaeota filo. Lähde: Liné1, CC BY-SA 3.0, Wikimedia Commons Tämä reuna määritettiin suhteellisen äskettäin, ja hyvin harvat siihen kuuluvista lajeista on viljelty In vitro, Näistä organismeista tiedetään niin vähän.
Kaikki Edge -jäsenet saavat energiansa ammoniumhapetuksesta ja jakautuvat maailmanlaajuisesti makean veden kappaleisiin, maaperiin, sedimenteihin ja kuumiin jousiin.
Filos Korachaeota, Aigachaeota ja Garchaeota
 Korachaeota
Korachaeota Jotkut alan asiantuntija -tutkijat, jotka perustuvat genomisekvenssien analysointiin, ovat äskettäin määrittäneet kolmen ylimääräisen reunan olemassaolon Archaea -valtakunnassa, vaikka näille reunoille ehdotetut lajit eivät vielä eristetty laboratoriossa.
Yleensä näihin reunoille kuuluvat jäsenet on löydetty monien maa- ja meriekosysteemien pinnan alla, mutta myös lämpövesillä ja syvänmeren hydrotermisissä järjestelmissä.
Ravitsemus
Suurin osa Chimiórophos -organismien kanssa varustetuista archaean -lajeista pystyy käyttämään erittäin vähentyneitä epäorgaanisia yhdisteitä saadakseen energiaa, jota he tarvitsevat "liikuttaa" metabolisia koneitaan, etenkin sellaisen, joka liittyy hengityksen kanssa.
Epäorgaanisten molekyylien "spesifisyys", joita he käyttävät substraattina energian tuottamiseksi, riippuu ympäristöstä, jossa kukin tietty laji kehittyy.
Muut kaarit, samoin kuin kasvit, levät, briofyyttit ja syanobakteerit, kykenevät suorittamaan fotosynteesiä, ts. He käyttävät ja muuntamaan aurinkosäteiden valonergian hyödylliseksi kemialliseksi energiaksi.
Se voi palvella sinua: proteiiniaminohapotOn osoitettu, että jotkut kaarit elävät joidenkin märehtijöiden vatsassa (pötsissä) (joista joukossa on lehmiä, lampaita, vuohia jne.), joten näitä kuvataan "keskinäisiksi kaareiksi", koska ne kuluttavat osan kuidusta, jota nämä eläimet syövät ja tekevät yhteistyötä joidenkin komponenttien ruuansulatuksen kanssa.
Jäljentäminen
Aivan kuten bakteerit, kaarit ovat yksisoluisia organismeja, joiden lisääntyminen on yksinomaan aseksuaalista. Lajista kuvatut tärkeimmät mekanismit In vitro ovat:
- Binaarinen fissio, jossa kukin arkki on "osa" puoliksi, joka on peräisin kahdesta identtisestä solusta
- Geminaatio tai "pirstoutuminen", jossa solut luovat "fragmentit" tai "osat" itsestään, jotka kykenevät muodostamaan uusia, geneettisesti identtisiä soluja.
Elinympäristö
Kaarit ovat pääosin liittyneet "äärimmäisiin" ympäristöihin, toisin sanoen niihin luonnollisiin paikkoihin, jotka asettavat vakavia rajoituksia elävien olentojen normaalille kehitykselle, etenkin lämpötilan, pH: n, suolapitoisuuden, anaerobioosin (hapen puuttumisen) suhteen jne.; Syy siihen, miksi opiskelusi on erittäin mielenkiintoinen, koska niillä on ainutlaatuiset mukautukset.
Viimeisimmät molekyylianalyysitekniikat kuitenkin käyttämättömien mikro -organismien lajien tunnistamiseksi (eristetty ja ylläpidetty In vitro Laboratoriossa) he ovat antaneet havaita kaarin esiintymisen.
Suurin osa luonnossa tunnistetuista kaareista luokitellaan kuitenkin heidän miehittämänsä elinympäristön mukaan, koska ne ovat kirjallisuuden sukulaisia termit "hypertermofiilit", "happofiilit" ja "äärimmäiset termasidofiilit", "äärimmäiset halofiilit" ja "metanogenos".
Hypertermofiilikaarien käyttämät ympäristöt ovat olemassa.
Ympäristöt, joissa äärimmäiset happofiilit elävät, ovat toisaalta niitä, joissa pH on erittäin alhainen, ja ne voidaan erottaa myös korkeilla lämpötiloissa (äärimmäiset termasidofiilit), sillä välin äärimmäisten halofiilien ympäristöt ovat ympäristöissä, joissa suolojen pitoisuus on erittäin korkea.
Metanogeeniset kaaret elävät ilman happea tai anaerobioosia, ympäristöissä, joissa voidaan käyttää muita molekyylejä, kuten elektronien vastaanottajia aineenvaihdunnassa, ja kykenevät tuottamaan metaania aineenvaihdunnan "jätteiden" tuotteena.
Esimerkkejä kaarilajeista
Tunnetut kaarilajeja on lukuisia, mutta tässä vain osa niistä tehdään.
Ignicoccus Hospitalis ja Nanoarchaeum Equitans
 Ignicoccus Hospitalis
Ignicoccus Hospitalis Yllyttää. Sairaala Se kuuluu Ignicoccus -nimisen Crenarqueotasin suvulle ja on quimiolitrofinen organismi, joka käyttää molekyylisen vety elektronien luovuttajana rikin pelkistykseen. Tällä lajilla on pienin genomi kaikista tähän mennessä kuvattuista kaareista.
Yllyttää. Sairaala Se käyttäytyy toisen lajin "loisena" tai "symbiootina": Nanoarchaeum Equitans. Jälkimmäistä ei ole viljelty In vitro Ja sen genomi on pienin kaikista kuvatuista kaareista, jotka on kuvattu.
Se asuu pääasiassa meriympäristöissä, eikä siinä ole geenejä lipidien, aminohappojen, nukleotidien tai kofaktorien biosynteesiin, joten kokeellinen näyttö viittaa siihen, että se saa nämä molekyylit vuorovaikutuksensa ansiosta sen kanssa Yllyttää. Sairaala.
Happolobus saccharovorans
Tämä on eräänlainen anaerobinen termasidofiilinen arkki, toisin sanoen, joka elää köyhissä tai täysin puutteellisissa happiympäristöissä, korkeat lämpötilat ja pH erittäin matala. Se löydettiin ensimmäistä kertaa Kamchatkan maanpäällisten kuumien lähteiden ruumiista.
Staphylothermus hellenicus
Tämä kaari kuuluu Crenarqueotasin reunaan, erityisesti desulfurokokin järjestykseen. Se on hypertermofiilinen heterotrofinen kaari (elää erittäin kuumissa ympäristöissä) ja vaatii rikkiä energian saamiseksi.