

Restriktioentsyymit, tyypit ja esimerkit
- 1119
- 68
- Sheldon Kuhn
Se rajoitusentsyymit Ne ovat endonukleaaseita, joita tietyt kaarit ja bakteerit käyttävät estämään tai "rajoittamaan" virusten etenemistä sisällä. Ne ovat erityisen yleisiä bakteereissa ja ovat osa heidän ulkomaisia DNA -puolustusjärjestelmäänsä, joka tunnetaan nimellä rajoitus-/modifikaatiojärjestelmä.
Nämä entsyymit katalysoivat kaksinkertaisen DNA: n leikkauksen tietyissä paikoissa, toistettavissa ja ilman lisäenergian käyttöä. Useimmat vaativat kofaktorien, kuten magnesiumin tai muiden kaksiarvoisten kationien, läsnäolon, vaikka jotkut vaativat myös ATP: tä tai S-adenosyylitationista.
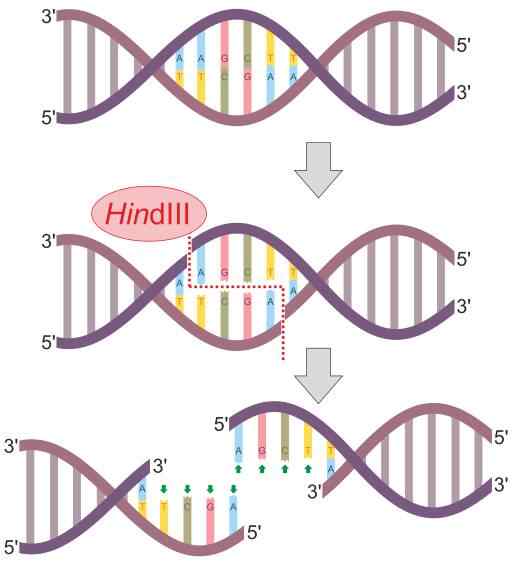
Hindii-restriktioentsyymireaktiokaavio (lähde: Helixitta [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)] Wikimedia Commonsin kautta) Daniel Nathans, Arber Werner ja Hamilton Smith löysivät vuonna 1978, jotka saivat Nobel -palkinnon lääketieteestä heidän löytöistä. Hänen nimensä on yleensä peräisin organismista, jossa niitä ensin havaitaan.
Tällaisia entsyymejä käytetään laajasti DNA -kloonausmenetelmien ja muiden molekyylibiologian ja geenitekniikan strategioiden kehittämisessä. Sen ominaispiirteet spesifisten sekvenssien tunnistamisessa ja sekvenssien leikkauskapasiteetissa tekevät niistä tehokkaita työkaluja geneettisessä kokeilussa.
Restriktioentsyymien tuottamia fragmentteja, jotka ovat vaikuttaneet tiettyyn DNA -molekyyliin.
Joillakin restriktioentsyymeillä voi olla sama tunnistuspaikka DNA: ssa, mutta ne eivät välttämättä leikkaa sitä samalla tavalla. Siten on entsyymejä, jotka tekevät leikkauksista jättäen ROMO: t ja entsyymit, jotka leikkaavat yhtenäisiä ääripäätä, joilla on erilaisia sovelluksia molekyylibiologiassa.
Kaupallisesti saatavissa on tällä hetkellä satoja erilaisia rajoitusentsyymejä, joita tarjoavat eri kaupalliset talot; Nämä entsyymit toimivat "räätälöityinä" molekyylisissä saksina eri tarkoituksiin.
[TOC]
Funktiot
Restriktioentsyymit täyttävät polymeraasien päinvastaisen toiminnan, koska ne hydrolysoituu tai katkaisee esterisidoksen fosfodiéster -sidoksessa vierekkäisten nukleotidien välillä nukleotidiketjussa.
Molekyylibiologiassa ja geenitekniikassa niitä käytetään laajalti työkaluja ekspression ja kloonausvektoreiden rakentamiseen sekä spesifisten sekvenssien tunnistamiseen. Ne ovat hyödyllisiä myös rekombinanttien genomien rakentamisessa ja niillä on suuri bioteknologinen potentiaali.
Geeniterapian viimeaikaiset edistysaskeleet hyödyntävät nykyistä restriktioentsyymejä vektoreissa määritettyjen geenien käyttöönottoon, jotka ovat ajoneuvoja tällaisten geenien kuljettamiseksi eläviin soluihin ja että heillä on todennäköisesti kyky työntää itsensä solun genomiin pysyvien muutosten suorittamiseksi.
Toimintamekanismi
Restriktioentsyymit voivat katalysoida kaksinauhan DNA -leikkausta, vaikka jotkut kykenevät tunnistamaan yksinkertaiset kaista -DNA -sekvenssit ja jopa RNA: n. Leikkaus tapahtuu sekvenssien tunnistamisen jälkeen.
Voi palvella sinua: glioksylaattisykli: Ominaisuudet, reaktiot, säätely, toiminnotVaikutusmekanismi koostuu fosfodiéster -yhteyden hydrolyysistä fosfaattiryhmän ja deoksiriboosin välillä kunkin DNA: n DNA: n luurankoon. Monet entsyymeistä kykenevät leikkaamaan samaan paikkaan, jonka he tunnistavat, kun taas toiset leikkaavat 5 - 9 emäsparia ennen tai jälkeen.
Normaalisti nämä entsyymit leikataan fosfaattiryhmän 5 'päässä, mikä johtaa DNA -fragmentteihin 5' fosforyylipäällä ja hydroksyylipäällä 3 'terminaalilla.
Koska proteiinit eivät ole suorassa kosketuksessa DNA: n tunnistuskohtaan, nämä on siirrettävä peräkkäisiä aikoja, kunnes erityinen paikka saavutetaan, ehkä DNA -juosteen "liukuvien" mekanismien avulla.
Entsymaattisen leikkauksen aikana kunkin DNA -juosteen fosfodiéster -linkki on sijoitettu johonkin aktiiviseen restriktioentsyymien kohtiin. Kun entsyymi poistuu tunnistus- ja leikkauskohdasta, se tekee niin epäspesifisten siirtymäyhdistysten kautta.
Kaverit
Tällä hetkellä tiedetään viisi restriktioentsyymejä. Seuraavaksi lyhyt kuvaus jokaisesta:
Tyypin I rajoitusentsyymit
Nämä entsyymit ovat suuria pentameerisiä proteiineja, joissa on kolme alayksikköä, yksi rajoituksista, metylaatio ja toinen DNA -sekvenssien tunnistamiseksi. Nämä endonukleaasit ovat monitoimisia proteiineja, jotka kykenevät katalysoimaan restriktio- ja modifikaatioreaktioita, niissä on ATPASA ja myös DNA topoisomera -aktiivisuus.
Tämän tyyppiset entsyymit olivat ensimmäiset löydetyt endonukleaasit, ne puhdistettiin ensimmäistä kertaa 1960 -luvulla, ja siitä lähtien niitä on tutkittu suurella syvyydellä.
Tyypin I entsyymejä ei käytetä laajasti bioteknologisena työkaluna, koska leikkuukohta voi olla muuttuvalla etäisyydellä 1.000 perusparia tunnistuskohdasta, mikä tekee niistä epäluotettavia kokeellisen toistettavuuden suhteen.
Tyypin II rajoitusentsyymit
Ne ovat entsyymejä, jotka koostuvat homodimeereistä tai tetrameereistä, jotka leikkaavat DNA: n paikkoihin, jotka on määritelty välillä 4 - 8 bp pituudeltaan. Nämä leikkauskohdat ovat tyypillisesti palindromisia, ts. Ne tunnistavat sekvenssit, jotka luetaan samalla tavalla molemmissa suunnissa.
Monet bakteerien tyypin II restriktioentsyymit leikkaavat DNA: ta, kun ne tunnistavat vieraan luonteensa, koska sillä ei ole tyypillisiä modifikaatioita, jotka omalla DNA: lla pitäisi olla.
Nämä ovat yksinkertaisempia restriktioentsyymejä, koska ne eivät vaadi muuta kofaktoria kuin magnesiumia (MG+) DNA -sekvenssien tunnistamiseksi ja leikkaamiseksi.
Tyypin II restriktioentsyymien tarkkuus yksinkertaisten sekvenssien tunnistamisessa ja leikkaamisessa DNA: ssa tarkissa paikoissa tekee niistä yhden eniten käytettyjä ja välttämättömiä useimmissa molekyylibiologian oksissa.
Voi palvella sinua: keskinäisyys: ominaisuudet, tyypit, esimerkitTyypin II restriktioentsyymien ryhmässä on useita alaluokkia, jotka luokitellaan tiettyjen ominaisuuksien mukaan, jotka ovat ainutlaatuisia jokaiselle. Näiden entsyymien luokittelu suoritetaan lisäämällä aakkosten kirjaimet A: sta z: hen entsyymin nimen jälkeen.
Jotkut niiden hyödyllisyyden tunnetuimmista alaluokista ovat:
Alaluokka IIA
Ne ovat erilaisia alayksikköä Dímeros. Ne tunnistavat epäsymmetriset sekvenssit ja niitä käytetään ihanteellisina esiasteina leikkausentsyymien muodostumisessa.
Alaluokka IIB
Ne koostuvat vielä yhdestä dimeeristä ja leikkaavat DNA: n tunnistussekvenssin molemmille puolille. He leikkasivat molemmat DNA -juosteet pohjaparien väliin myöhemmin tunnistuspaikasta.
IIC alaluokka
Tämän tyyppiset entsyymit ovat polypeptidejä DNA -juosteiden jakautumis- ja modifikaatiofunktioilla. Nämä entsyymit leikkaavat molemmat säikeet epäsymmetrisesti.
Alaluokka iie
Tämän alaluokan entsyymit ovat eniten käytettyjä geenitekniikassa. Heillä on katalyyttinen paikka ja yleensä ne vaativat alestron efektoria. Näiden entsyymien on oltava vuorovaikutuksessa niiden tunnistussekvenssin kahden kopion kanssa tehokkaan leikkauksen tekemiseksi. Tämän alaluokan sisällä ovat ECORII- ja ECORI -entsyymejä.
Tyypin III rajoitusentsyymit
Tyypin III rajoitus Endonukleaasit koostuvat vain kahdesta alayksiköstä, toinen vastaa DNA: n tunnistamisesta ja modifioinnista, kun taas toinen on vastuussa sekvenssin leikkauksesta.
Nämä entsyymit vaativat kaksi kofaktoria niiden toimintaan: ATP ja magnesium. Tämän tyyppisillä restriktioentsyymeillä on kaksi epäsymmetristä tunnistuskohtaa, ne siirtävät DNA: n ATP -riippuvaisella tavalla ja leikkaa se välillä 20–30 bp tunnistuskohdan vieressä.
Tyypin IV rajoitusentsyymit
Tyypin IV entsyymejä on helppo tunnistaa, koska ne leikkaavat DNA: n metylaatiomerkeillä, ne koostuvat useista eri alayksiköistä, jotka ovat vastuussa DNA -sekvenssin tunnistamisesta ja leikkaamisesta. Nämä entsyymit käyttävät GTP -kofaktoreita ja kalliita magnesiumia.
Spesifisiä leikkauskohtia ovat nukleotidiketjut, joissa metyloituja tai hydroksymetyleitä sytosiinitähteitä yhdessä tai molemmissa nukleiinihappojuoksissa.
Tyypin V rajoitusentsyymit
Tämä luokitusryhmät Crisper-Cas-tyyppiset entsyymit, jotka tunnistavat ja leikkaavat tunkeutuvien organismien spesifiset DNA-sekvenssit. CAS -entsyymit käyttävät RNA -juosteen syntetisoitua opasta Crisperille tunnistamaan ja hyökkäämään tunkeutuviin organismeihin.
Tyypiksi V luokitetut entsyymit ovat tyypin I, II ja II entsyymien rakenteellisia polypeptidejä. Ne voivat leikata melkein minkä tahansa organismin DNA -osia ja suurella pituudella. Sen joustavuus ja helppo työllisyys tekevät näistä entsyymeistä yhden geenitekniikan eniten käytetyistä työkaluista yhdessä tyypin II entsyymien kanssa.
Voi palvella sinua: PropionibacteriumEsimerkit
Restriktioentsyymejä on käytetty DNA -polymorfismien havaitsemiseen, etenkin populaatiogenetiikan tutkimuksissa ja evoluutiotutkimuksissa, joissa käytetään mitokondriaalista DNA: ta, nukleotidin substituutioasteen tiedon saamiseksi.
Tällä hetkellä bakteerien muuntamiseen käytettyjen vektorien kanssa eri tarkoituksiin on monikerroksisia kohtia, joissa löydetään useita restriktioentsyymejä varten tunnistuskohtia.
Näiden entsyymien joukossa suosituimpia ovat ECORI, II, III, IV ja V, saatu ja kuvattu ensimmäistä kertaa JA. koli; Hindiii, alkaen H. influenza ja bamhi B -. Amyyloliquefaciens.
Viitteet
- Bikle, t. -Lla., & Kruger, D. H. (1993). DNA -rajoituksen biologia. Mikrobiologiset arvostelut, 57(2), 434-450.
- Boyval, s., Moineau, S., Romero, D. -Lla., & Horvath, P. (2007). CRISPR antaa vakuutuksen Prokaryootien virusten suhteen. Tiede, 315(Maaliskuu), 1709-1713.
- Goodsell, D. (2002). Molekyylin näkökulma: rajoitus endonukleaasit. Kantasolut syöpälääketieteen perusteet, kaksikymmentä, 190-191.
- Puoliard, s. JA. (2001). Hyppääminen, hyppääminen ja silmukointi restriktioentsyymeillä. Biokemialliset seuran liiketoimet, 29, 363-373.
- Jeltsch, a. (2003). Lajien identiteetin ylläpitäminen ja bakteerien spesifikaatio: uusi funktio rajoitus-/modifikaatiojärjestelmille? Geeni, 317, 13-16.
- Krebs, J., Goldstein, E., & Kilpatrick, S. (2018). Lewinin geenit xii (12 Ed.-A. Burlington, Massachusetts: Jones & Bartlett Learning.
- Li, ja., S, s., Zhang, ja., Ren, m., Feng, m., Peng, n.,... hän, Q. (2015). Tyypin I ja tyypin III CRISPR-CAS-järjestelmien valjaus genomin muokkaamiseen. Nukleiinihappotutkimus, 1-12.
- Loenen, W. -Lla. M., Dryden, D. T. F., Raleight, E. -Lla., & Wilson, G. G. (2013). Tyypin I rajoitusentsyymit ja heidän sukulaiset. Nukleiinihappotutkimus, 1-25.
- Nathans, D., & Smith, H. JOMPIKUMPI. (1975). Restriktio -endonukleaasit DNA -molekyylien analysoinnissa ja uudelleenjärjestelyissä. Annu. Rev. Biokemia., 273-293.
- Nei, m., & Tajima, f. (1981). DNA -polymorfismi, joka voidaan havaita restriktio -endonukleaaseilla. Genetiikka, 145-163.
- Pingoud, a., Fuxreiter, m., Pingoud, v., & Wende, W. (2005). Solu- ja molekyylieläintieteiden tyypin II restriktio -endonukleaasit: rakenne ja mekanismi. CMLS -solu- ja molekyylieläintieteet, 62, 685-707.
- Roberts, r. (2005). Kuinka restriktioentsyymeistä tuli molekyylibiologian työhevoset. PNA: t, 102(17), 5905-5908.
- Roberts, r. J -., & Murray, k. (1976). Rajoitukset. Kriittiset arvostelut biokemiassa, (Marraskuu), 123-164.
- Stoddard, b. Lens. (2005). HOMING -endonukleaasirakenne ja toiminta. Biofysiikan neljännesvuosittaiset arvostelut, 1-47.
- Tock, m. R -., & Dryden, D. T. F. (2005). Rajoituksen ja lenkilön vastaisen biologia. Nykyinen mielipide mikrobiologiassa, 8, 466-472. https: // doi.org/10.1016/j.Minun b.2005.06.003
- Wilson, G. G., & Murray, n. JA. (1991). Rajoitus- ja muokkausjärjestelmät. Annu. Rev. Genetti., 25, 585-627.
- Wu, z., & MOU, K. (2016). Genomiset näkemykset Campylobacter jejuni -virulenssista ja populaation genetiikasta. Infec. Dis. Kääntää. Lääketieteellinen., 2(3), 109-19.
- Yuan, r. (1981). Monitoimisten rajoitusten rakenne ja mekanismi. Annu. Rev. Biokemia., viisikymmentä, 285-315.
- « Kirjailijat ja teokset 1914 -ominaisuuksien sukupolvi
- Toisiaan poissulkevia tapahtumien ominaisuuksia ja esimerkkejä »