

Sphingolipidit mitä ovat, ominaisuudet, toiminnot, synteesi
- 4019
- 337
- Kelly Kilback
Mitä ovat sphylingolipidit?
Se sfingolipidit Ne edustavat yhtä kolmesta suuresta lipidiperheestä, jotka ovat läsnä biologisissa kalvoissa. Kuten glysofosfolipidit ja sterolit, ne ovat amfipaattisia molekyylejä, joilla on hydrofiilinen polaarinen alue ja hydrofobinen apolaarinen alue.
Johann L kuvasi heidät ensimmäisen kerran vuonna 1884. W -. Thudichum, joka kuvasi kolmea sfingolipidiä (sfingomyeline, aivo- ja aivo -aivo), jotka kuuluvat kolmeen eri luokkaan, jotka tunnetaan: fosfosfingolipidit, glykosfingolipidit neutraalit ja happamat.
Toisin kuin glysofosfolipidejä, sphyyolipidejä ei ole rakennettu 3-fosfaattiglyserolimolekyylille pääluukkuksi, vaan ovat yhdisteitä, jotka ovat peräisin sfhinyltä, aminoalkoholista, jolla.
JASfingolipidien rakenne
Kaikki sfingolipidit johdetaan L-seriinistä, joka on tiivistetty pitkäketjuisella rasvahapolla sfingoidipohjan muodostamiseksi, joka tunnetaan myös nimellä pitkäketjuinen emäs (LCB).
Yleisimmät emäkset ovat sfinganiini ja sfinksiini, jotka eroavat toisistaan kaksoissidoksen läsnä ollessa trans Sfinksiinin rasvahapon 4 ja 5 välillä.
Sfinksiinin hiilet 1, 2 ja 3 ovat rakenteellisesti analogisia glyserolin glyserofosfolipidien kanssa glyseroli. Kun AMIDA -sidokset sitovat rasvahappoa hiili -2: een sfiiniä.
Näiden lipidien hydrofobiset alueet muodostavat pitkät ketjut rasvahapot voivat olla hyvin monimuotoisia. Pituudet vaihtelevat 14 - 22 hiiliatomia, joilla voi olla erilaiset kylläisyysasteet, yleensä hiilihiilien välillä 4 - 5.
Asemissa 4 tai 6 niillä voi olla hydroksilli -ryhmiä ja kaksoissidoksia muissa paikoissa tai jopa seuraamuksissa, kuten metyyliryhmät.
Ominaisuudet
Amida -sidosten yhdistämät rasvahappoketjut keramiidiin ovat yleensä kylläisiä, ja ne ovat yleensä suurempia kuin glyserofosfolipideissä, jotka näyttävät olevan ratkaisevia näiden biologiselle aktiivisuudelle.
Sphingolipid -luurankon erottuva ominaisuus on, että niillä voi olla positiivinen nettokuorma neutraaliin pH: han, outo lipidimolekyylien keskuudessa.
Kuitenkin PkAminoryhmän a on alhainen yksinkertaisen amiinin suhteen, välillä 7 - 8, joten osaa molekyylistä ei ladata fysiologiseen pH: han, mikä voisi selittää näiden "vapaan" liikkeen Bilapasin keskuudessa.
Sfingolipidien perinteinen luokittelu johtuu monista modifikaatioista, joita keramidimolekyyli voi kärsiä, etenkin polaaristen pääryhmien substituutioiden suhteen.
Voi palvella sinua: Hepadnavirus: Ominaisuudet, morfologia, hoitoFunktiot
Sfingolipidit ovat välttämättömiä eläimissä, kasveissa ja sienissä, samoin kuin joissain prokaryoottisissa organismeissa ja viruksissa.
-Rakennetoiminnot
Sfingolipidit moduloivat kalvojen fysikaalisia ominaisuuksia, mukaan lukien niiden juoksevuus, paksuus ja kaarevuus. Modulaariset nämä ominaisuudet antavat heille myös suoran vaikutuksen kalvoproteiinien alueelliseen organisointiin.
Lipidissä "lautat"
Biologisissa kalvoissa dynaamiset mikrodomeenit voidaan havaita alhaisemmalla sujuvuudella muodostettuna kolesterolilla ja sfingolipidimolekyyleillä, joita kutsutaan lipidibalsasiksi.
Nämä rakenteet esiintyvät luonnollisesti ja ylläpitävät läheistä suhdetta kattavien proteiinien, solujen pintareseptoreiden ja signalointiproteiinien, kuljettajien ja muun proteiinin kanssa glykosyylifosfatidyylinitoli (GPI) -ankkureiden (GPI) kanssa (GPI).
-Signalointitoiminnot
Heillä on toimintoja, kuten signalointimolekyylejä, jotka toimivat toisina lähettiläinä tai erittyneinä ligandeina solun pintareseptoreille.
Toissijaisina lähettiläinä he voivat osallistua kalsiumin homeostaasin, solujen kasvun, tuumorogeneesin ja apoptoosin tukahduttamisen säätelyyn. Lisäksi monien integraali- ja perifeeristen membraaniproteiinien aktiivisuus riippuu sen yhteydestä sfingolipidien kanssa.
Monet solujen väliset ja solujen vuorovaikutukset niiden ympäristön kanssa riippuvat sfingolipidien eri polaaristen ryhmien altistumisesta kohti plasmamembraanin ulkopintaa.
Glykosfingolipidien ja lektinien liitto on ratkaisevan tärkeää myeliinin assosiaatiolle aksonien kanssa, neutrofiilien tarttuminen endoteeliin jne.
Hänen aineenvaihdunnan tuotteet
Tärkeimmät signaloinnin sfingolipidit ovat pitkän ketjun emäkset tai sfinksit ja keramiidit, samoin kuin niiden fosforyloidut johdannaiset, kuten 1-fosfaatti-sphygosiini.
Monien sfingolipidien aineenvaihduntatuotteet aktivoivat tai estävät useita valkoisia ala -asiaa (kinaasiproteiineja, fosfatoosia ja muita), jotka hallitsevat solujen käyttäytymistä yhtä monimutkaisia kuin kasvu, erilaistuminen ja apoptoosi.
-Kalvoreseptoreina
Jotkut patogeenit käyttävät gloysfingolipidejä reseptoreina välittämään heidän sisäänkäynninsä isäntäsoluihin tai toimittamaan heille virulenssitekijöitä.
On osoitettu, että sfyglyipidit osallistuvat useisiin soluihin, kuten eritys, endosytoosi, kemiotaxis, neurotransmissio, angiogeneesi ja tulehdus.
Ne ovat mukana myös kalvoliikenteessä, joten ne vaikuttavat reseptorien internalisaatioon, erittyvien vesikkelien järjestyksen, liikkumisen ja fuusion vastauksena eri ärsykkeille.
Sfingolipidiryhmät
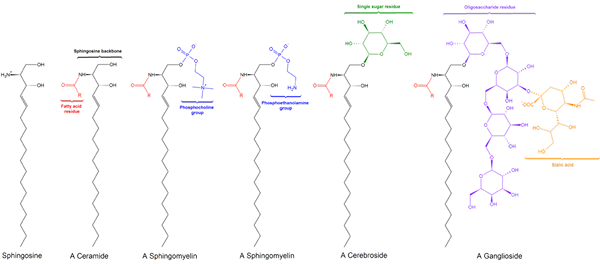
Sfhingolipid -alaluokkaa on kolme, jotka kaikki ovat peräisin keramidista ja jotka eroavat toisistaan polaariryhmillä, nimittäin: sphylingomyeliinit, glykolipidit ja gangliasidit.
Sfingomyielinas
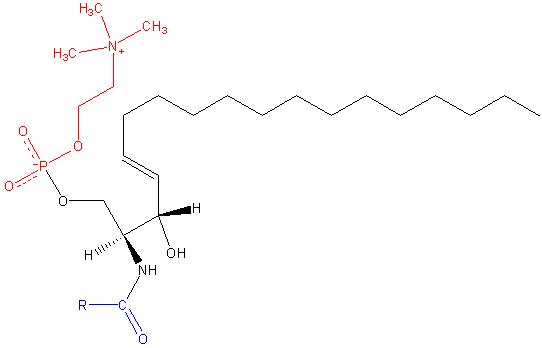 Sfhingomileiini. Musta: Sfinksiin. Punainen: fosfokoliini. Sininen: Rasvahappo.
Sfhingomileiini. Musta: Sfinksiin. Punainen: fosfokoliini. Sininen: Rasvahappo. Nämä sisältävät fosfokoliinia tai fosfoetanoliamiinia polaarisena pääryhmänä, joten ne luokitellaan fosfolipideiksi yhdessä glyceofosfolipideiksi. Ne näyttävät tietysti fosfatidyylikoliineilta kolmen dimensionaalisessa rakenteessa ja yleisominaisuuksissa, koska niillä ei ole veloitusta napapäässään.
Voi palvella sinua: enolaasi: rakenne, toimintamekanismi, toiminnotNe ovat läsnä eläinsolujen plasmamembraaneissa ja ovat erityisen runsaasti myeliinissä, podissa, joka ympäröi ja eristää joidenkin neuronien aksonit.
Glykolipidit tai neutraalit glykosfingolipidit (ilman kuormaa)
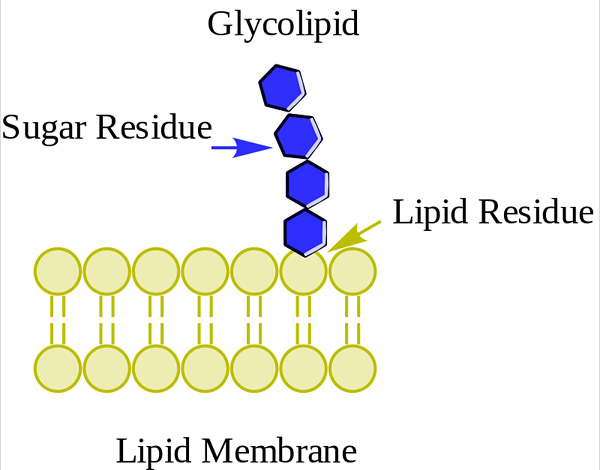 Glukolipidi. WPCROSSON [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons
Glukolipidi. WPCROSSON [CC BY-SA 4.0 (https: // creativecommons.Org/lisenssit/by-SA/4.0)], Wikimedia Commons Ne ovat ensisijaisesti plasmamembraanin ulkopinnassa ja niillä on yksi tai useampi sokeri napapääryhmänä, joka on suoraan yhdistynyt hiilihydroksyyli 1. Heillä ei ole fosfaattiryhmiä. Koska pH 7: llä ei ole kuormaa, niitä kutsutaan neutraaliksi glykolipideiksi.
Aivobrosidit ovat yksi sokerimolekyyli, joka on kytketty keramiidiin. Ne, jotka sisältävät galaktoosia. Globosidit ovat goustoesfingolipidejä, joissa on kaksi tai useampia sokereita, yleensä D-glykoosi, D-galaktoosi tai N-asetyyli-D-galaktosamiini.
Gangliósidos tai glykosfingolipidit
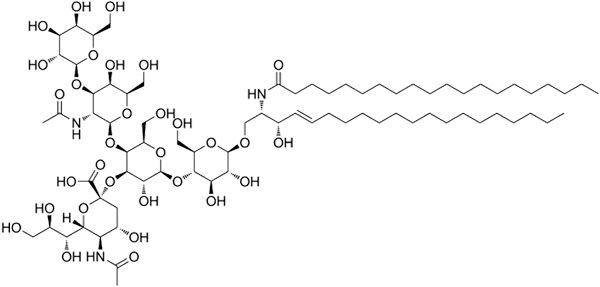 GM1 -ganglosidirakenne
GM1 -ganglosidirakenne Nämä ovat monimutkaisimpia sfingolipidejä. Heillä on oligosakkarideja polaarisena pääryhmänä ja yhtenä tai useamman terminaalisen N-asetyylimuramihappotähteenä, jota kutsutaan myös siralihapoksi. Siamiinihappo tuo gangliaan negatiivisen kuormituksen pH 7: een, joka erottaa ne neutraalista glykosfingolipideistä.
Tällaisten sfingolipidien nimikkeistö riippuu polaarisen pään oligosakkaridiosassa esiintyvän sisaliinihappojätteen määrästä.
Synteesi
Pitkän ketju- tai sphiny -emäsmolekyyli syntetisoidaan endoplasmisessa retikulumissa (ER) ja näiden lipidien päähän liittyvän polaariryhmän lisääminen tapahtuu myöhemmin Golgi -kompleksissa. Nisäkkäissä jonkin verran sfingolipidien synteesiä voi esiintyä myös mitokondrioissa.
Saatuaan synteesin Golgi -kompleksissa, sfingolipidit kuljetetaan muihin soluosastoihin vesikkelien välittämien mekanismien kautta.
Sfingolipidien biosynteesi koostuu kolmesta perustapahtumasta: pitkien ketjun emäksen synteesi, keramidien biosynteesi rasvahapon liitoksella Amida -sidoksen kautta ja lopuksi kompleksin sfingolipidien muodostuminen unionin avulla. polaariryhmiä hiili 1 sfingoidepohjan.
Synteesin lisäksi Novo, Sfingolipidit voivat muodostua myös pitkien ketjujen ja keramiidien korvaamisella tai kierrättämällä, jotka voivat ruokkia sfingolipidiallasta.
Keramidin luurankon synteesi
Keramidin biosynteesi, sfingolipidien luuranko, alkaa palmitail-CoA-molekyylin ja L-seriinin dekarboksyleratiivisella tiivistymisellä. Reaktiota katalysoi seriinipalmitail-transferaasi (SPT) heterodimeerinen, pyridoksaalinen fosfaattiriippuva ja tuote on 3-Zo dihydroesfingosiini ze.
Voi palvella sinua: mikrobiota: koostumus, toiminnot ja missä se onTätä entsyymiä estävät β-phaal-l-alaniinit ja L-Cicloserinas. Hiivoissa se koodaa kaksi geeniä, kun taas nisäkkäissä on kolme geeniä tälle entsyymille. Aktiivinen kohta on endoplasmisen retikulumin sytoplasmisessa pinnassa.
Tämän ensimmäisen entsyymin rooli säilytetään kaikissa tutkituissa organismeissa. Taksonien välillä on kuitenkin joitain eroja, jotka liittyvät entsyymin solun sijainnin kanssa: bakteerit ovat sytoplasmista, hiiva, kasvit ja eläimet ovat endoplasmisessa retikulumissa.
3-zoesfinganiini vähenee myöhemmin NADPH-riippuvalla 3-Zo-ketofinganiinilla. Dihydrokeramidisyntaasi (sfinganiini N-akilin transferaasi)-asetyyli sitten sfinaniini dihydrokeramidin tuottamiseksi. Sitten keramidi muodostuu dihydrokeramidi -deaturaasi/reduktaasi, joka lisää kaksoissidoksen trans Paikoillaan 4-5.
Nisäkkäissä on lukuisia keramidisomformeja syntasas, jokainen yhdistyy tiettyyn rasvahappoketjuun pitkien ketjujen emäksiin. Siksi keramidit synteesat ja muut entsyymit, elongasat, tarjoavat tärkeimmän rasvahappojen monimuotoisuuden lähteen sfingolipideissä.
Aineenvaihdunta
Sfingolipidien hajoaminen suorittavat glykolikohydrolit ja sfingomyelinaasientsyymit, jotka ovat vastuussa polaaristen ryhmien modifikaatioiden poistamisesta. Toisaalta keramiidit uudistavat pitkät ketjut emäkset keramiidista.
Gangliat hajoavat sarjassa lysosomaalisia entsyymejä, jotka katalysoivat sokeriyksiköiden askel askeleelta, tuottaen keramidin lopulta.
Toinen hajoamisvapa on sfingolipidien internalisointi endosyyttisissä vesikkeleissä, jotka lähetetään takaisin plasmamembraaniin tai kuljetetaan lysosomeihin, joissa ne hajoavat spesifisen happorydrolaasin avulla.
Kaikkia pitkän ketjun emäksiä ei kierrätetä, endoplasminen säilyy. Tämä hajoamismekanismi koostuu fosforylaatiosta LCB: ien käytön sijasta, mikä johtaa signalointimolekyyleihin, jotka voivat olla liukoisia substraatteja valhesentsyymeille.
Säätö
Näiden lipidien aineenvaihduntaa säädetään useilla tasoilla, yksi niistä on synteesin, niiden jälkeisten translaation jälkeisten modifikaatioiden ja samojen mekanismien, entsyymit ja samat mekanismit.
Jotkut säätelymekanismit ovat solu-spesifikaatioita, joko hallita solujen kehitysaikaa, jossa ne tuotetaan, tai vasteena erityisiin signaaleihin.
Viitteet
- Bartke, n., & Hannun, ja. (2009). Bioaktiiviset spfingolipidit: aineenvaihdunta ja toiminta. Journal of Lipid Research, viisikymmentä, 19.
- Breslow, D. K -k -. (2013). Sfingolipidi homeostaasi endoplasmisessa retikulumissa ja sen ulkopuolella. Cold Spring Harbor -näkymät biologiassa, 5(4), A013326.