

Nukleosomi
- 701
- 75
- Louis Moen
Nukleosomi on DNA: n organisaation perustavanlaatuinen yksikkö eukaryoottisoluissa Mikä on nukleosomi?
Hän Nukleosomi Se on eukaryoottisten organismien DNA -pakkausyksikkö. Siksi se on pienin kromatiinin puristuselementti.
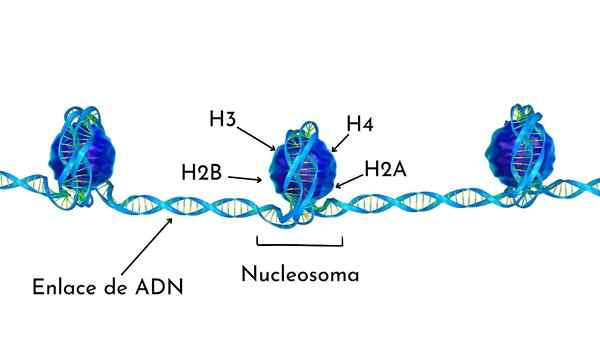
Nukleosomi on rakennettu proteiinioktameeriksi, nimeltään histonit, tai rummun muotoinen rakenne, jolla on noin 140 nt DNA: ta, mikä antaa melkein kaksi täydellistä kierrosta, kuten kuvassa voidaan nähdä.
Katsottaan, että jotkut ylimääräiset DNA -NT: t ovat osa nukleosomia ja on DNA -fraktio, joka sallii fysikaalisen jatkuvuuden yhden nukleosomin ja toisen välillä monimutkaisemmissa kromatiinirakenteissa (kuten kromatiinikuitu 30 nm).
Histonikoodi oli yksi ensimmäisistä paremmista epigeneettisistä ohjauselementeistä, jotka ymmärrettiin molekyylisesti.
Nukleosomi toimii
Nukleosomit sallivat:
- DNA -pakkaukset rajoitetun ydinalueen sijoittamiseksi.
- Ne määrittelevät ekspressoituneen kromatiinin (euchromatiinin) ja hiljaisen kromatiinin (heterokromatiinin) väliseudun jakautumisen välillä.
- Järjestä kaikki kromatiinit sekä spatiaalisesti että toiminnallisesti ytimessä.
- Ne edustavat kovalenttisten modifikaatioiden substraattia, jotka määrittävät proteiinien koodauksen ekspression ja ekspression tason, niin soittaman histonikoodin kautta.
Koostumus ja rakenne
Perusteellisimmassa mielessä nukleosomit koostuvat DNA: sta ja proteiineista. DNA voi olla käytännöllisesti katsoen mikä tahansa kaksikaistainen DNA, joka on läsnä eukaryoottisen solun ytimessä, kun taas kaikki nukleosomaaliset proteiinit kuuluvat proteiinisarjoihin, joita kutsutaan histoniksi.
Voi palvella sinua: solujen erotteluHistonit ovat pieniä proteiineja ja suurella aminohappojätteen kuormituksella, joka mahdollistaa DNA: n korkean negatiivisen kuormituksen torjumisen ja muodostaa tehokkaan fysikaalisen vuorovaikutuksen kahden molekyylin välillä saavuttamatta kovalenttisen kemiallisen sidoksen jäykkyyttä.
Histonit muodostavat rumpu -tie oktametrin, jossa on kaksi kopiota tai monomeeriä jokaisesta H2A: sta, H2B-, H3- ja H4 -histonista.
DNA antaa melkein kaksi täydellistä käännöstä oktaamerin sivuilla ja jatkuu sitten murto -osalla linkker -DNA: ta, joka liittyy histoni H1: hen, palatakseen kahteen täydelliseen käännökseen toisessa Histona Octa -sovelluksessa.
Oktametrisarja, siihen liittyvä DNA ja sen vastaava linkkeri -DNA on nukleosomi.
 Nukleosomin osat
Nukleosomin osat Kromatiinin tiivistys
Genominen DNA muodostuu erittäin pitkistä molekyyleistä (enemmän kuin yksi metri ihmisen tapauksessa, ottaen huomioon kaikki sen kromosomit), joiden on oltava yhteensopivia ja organisoituneita erittäin pienessä ytimessä.
Tämän tiivistyksen ensimmäinen vaihe suoritetaan nukleosoomien muodostumisen kautta. Vain tällä vaiheella DNA on tiivistetty noin 75 kertaa.
Tämä aiheuttaa lineaarisen kuidun, josta seuraavat kromatiinin tiivistystasot on rakennettu: 30 nm: n kuitu, siteet ja siteet siteet.
Kun solu on jaettu joko mitoosilla tai meioosilla, viimeinen tiivistysluokka on mitoottinen tai meioottinen kromosomi itse, vastaavasti itse.
Histonikoodi ja geneettinen ekspressio
Se tosiasia, että histoni oktameerit ja DNA ovat vuorovaikutuksessa sähköstaattisesti selittää sen tehokkaan assosiaation menettämättä nukleosomien dynaamisia tiivistymiselementtejä ja kromatiinin dynaamista.
Voi palvella sinua: solujen hajoaminenMutta on vielä yllättävämpi vuorovaikutuselementti: Histonien päät Nitteet paljastetaan oktametrin sisäpuolelle, kompakti ja inertti.
Nämä äärimmäisyydet eivät ole vain vuorovaikutuksessa fyysisesti DNA: n kanssa, vaan kärsivät myös sarjan kovalenttisia modifikaatioita, joista kromatiinin tiivistymisaste ja siihen liittyvä DNA: n ekspressio riippuvat.
Kovalenttisten modifikaatioiden joukko tyypin ja numeron suhteen muun muassa tunnetaan kollektiivisesti nimellä Histon -koodi.
Nämä modifikaatiot sisältävät fosforylaation, metylaation, asetyloinnin, ubikvitinaation ja voimakkaasti.
Jokainen muutos määrittää liittyvän DNA: n ekspression tai ei, samoin kuin kromatiinin tiivistymisaste yhdessä muiden muutosten kanssa saman molekyylin sisällä tai muiden histonien, erityisesti H3 H3: n jätteiden kanssa.
Yleisesti ottaen on nähty esimerkiksi, että hypermetiloituneet ja hypakloituneet histonit määrittävät, että siihen liittyvää DNA: ta ei ekspressoitu ja että kromatiini esitetään kompaktimmassa tilassa (heterokromaattinen ja siten inaktiivinen).
Päinvastoin, eukromaattinen DNA (vähemmän kompakti ja geneettisesti aktiivinen) liittyy kromatiiniin, jonka histonit ovat hyperistetiiloituja ja hypometylisoituja.
Euchromatiini ja heterokromatiini
Histonasin kovalenttinen modifikaatiotila voi määrittää paikallisen kromatiinin ilmentymisasteen ja tiivistymisen.
Globaaleilla tasoilla kromatiinin tiivistymistä säätelevät tasaisesti nukleosomien histonien kovalenttiset modifikaatiot.
Esimerkiksi on osoitettu, että konstitutiivinen heterokromatiini (jota ei koskaan ilmaista ja tiheästi pakattu) on yleensä kiinnitetty ydinlevyyn, jättäen ydinhuokoset vapaiksi.
Voi palvella sinua: kalsiumpumppu: toiminnot, tyypit, rakenne ja toimintaKonstitutiivinen euchromatiini puolestaan (joka on aina ekspressoitu, kuten sellainen, joka sisältää solujen ylläpitogeenejä ja joka sijaitsee laiskromatiinialueilla), tekee niin suurissa siteissä, jotka paljastavat DNA: n transkriptiokoneeseen.
Muut genomisen DNA: n alueet värähtelevät näiden kahden tilan välillä riippuen organismin kehitysajasta, kasvuolosuhteista, soluidentiteetistä jne.
Muut toiminnot
Solujen kehitys-, ekspressio- ja ylläpitosuunnitelmansa toteuttamiseksi eukaryoottisten organismien genomien on hienosti säädettävä milloin ja miten niiden geneettiset potentiaalit tulisi ilmetä.
Alkaen geeneihin tallennetuista tiedoista, nämä sijaitsevat ytimessä yksityisillä alueilla, jotka määrittävät niiden transkription tilan.
Siksi voimme sanoa, että toinen nukleosoomien peruspapereista on niiden ytimen organisaatio tai arkkitehtuuri.
Tämä arkkitehtuuri on peritty ja fylogeneettisesti säilynyt näiden informaatiopakkausten modulaaristen elementtien olemassaolon ansiosta.
Viitteet
- Brooker, r. J -. Genetiikka: analyysi ja periaatteet. McGraw-Hillin korkeakoulu.
- Goodenough, u. W -. Genetiikka. W -. B -. Saunders Co. OY.