

Polyploidiatyypit eläimissä, ihmisissä, kasveissa
- 2973
- 288
- Mr. Clifford Kshlerin
Se Polyploidia Se on eräänlainen geneettinen mutaatio, joka koostuu kromosomien täydellisen rahaston (täydellisten pelien) lisäämisestä solun ytimeen, joka muodostaa homologisia ikätovereita. Tämäntyyppinen kromosomaalinen mutaatio on yleisin euploidioista, ja sille on ominaista organismilla on kolme tai useampia täydellisiä kromosomeja pelejä.
Organismia (diploidi normaalisti = 2n) pidetään polyploidina, kun se hankkii yhden tai useamman täydellisen kromosomisarjan. Toisin kuin spesifiset mutaatiot, kromosomaaliset sijoitukset ja päällekkäisyydet, tämä prosessi on laajamittainen, ts. Se tapahtuu täydellisissä kromosomeissa.
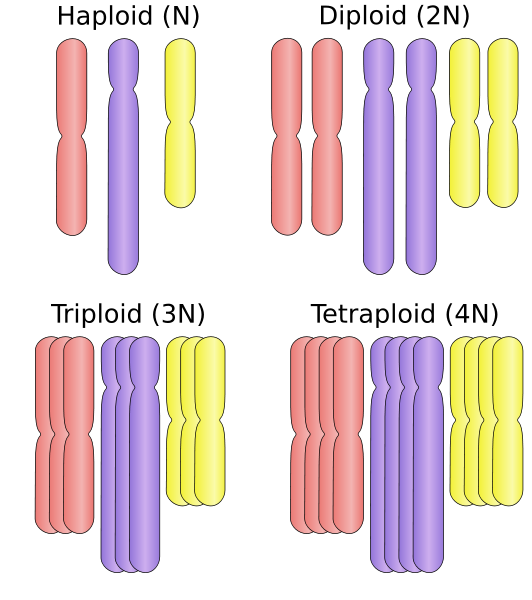
Lähde: HAPLOID_VS_DIPLOID.SVG: Ehambergderiivinen työ: Ehamberg [CC BY-SA 3.0 (https: // creativecommons.Org/lisenssit/by-SA/3.0)] Sen sijaan, että se olisi haploidinen (N) tai diploidi (2N), polyploidiorganismi voi olla tetraploidi (4N), oktoploidi (8N) tai suurempi. Tämä mutaatioprosessi on melko usein kasveissa ja on outo eläimissä. Tämä mekanismi voi lisätä geneettistä variaatiota niissä istunnissa, jotka eivät pysty siirtymään ympäristöstä.
Polyploidialla on suuri merkitys evoluutiotermeissä tietyissä biologisissa ryhmissä, joissa se muodostaa usein uusien lajien tuottamisen mekanismia, koska kromosomaalinen kuorma on perinnöllinen tila.
[TOC]
Kun polyploidia tapahtuu?
Kromosomien lukumäärän häiriöt voivat tapahtua sekä luonnossa että laboratoriossa vahvistetuissa populaatioissa. Ne voidaan indusoida myös mutageenisella aineella, kuten matot. Meioosin uskomattomasta tarkkuudesta huolimatta kromosomaalisia poikkeavuuksia tapahtuu ja ovat yleisempiä kuin voi ajatella.
Polyploidia syntyy joidenkin muutosten seurauksena, joita voi tapahtua meioosin aikana joko ensimmäisessä meioottisessa jakautumisessa tai profaasin aikana, joissa homologiset kromosomit on järjestetty pareittain tetradien muodostamiseksi ja jälkimmäisten aikana tapahtuva dishyklict Anafaasi I.
Uusien lajien esiintyminen
Polyploidia on tärkeä, koska se on lähtökohta uusille lajeille. Tämä ilmiö on tärkeä geneettisen variaation lähde, koska se aiheuttaa satoja tai tuhansia kaksoiskappaleita, jotka voivat vapaasti saada uusia toimintoja.
Kasveissa se on erityisen tärkeä ja melko laajalle levinnyt. On arvioitu, että yli 50% kukkasaiista on peräisin polyploidiasta.
Useimmissa tapauksissa polyploidit eroavat fysiologisesti alkuperäisistä lajeista ja tästä johtuen ne voivat kolonisoida ympäristöjä, joilla on uusia ominaisuuksia. Monet tärkeät lajit maataloudessa (mukaan lukien vehnä) ovat hybridi -alkuperän polyploideja.
Polyploidiatyypit
Polyloidia voidaan luokitella solun ytimessä olevien kokonaisjoukkojen tai kromosomaalisten pelien lukumäärän mukaan.
Tässä mielessä organismi, joka sisältää ”kolme” kromosomia, on “triploidi”, “tetraploidi”, jos se sisältää 4 kromosomia, pentaploidia (5 peliä), heksaploidae (6 peliä), heptoploidia (seitsemän peliä), oktoploidia (kahdeksan peliä), Nonaploidae (yhdeksän peliä), estäminen (10 peliä) ja niin edelleen.
Voi palvella sinua: ekologinen eristys: mikä on, mekanismi ja esimerkitToisaalta polyploidiat voidaan myös luokitella kromosomaalisten rahoitusten alkuperän mukaan. Tässä ideojärjestyksessä organismi voi olla: autopoliploidi tai alopoliploidi.
Autopoliploidi sisältää useita homologisten kromosomien pelejä, jotka ovat peräisin samasta yksilöstä tai samasta lajista kuuluva henkilö. Tässä tapauksessa polyploidit muodostuvat yhdistämällä geneettisesti yhteensopivien organismien vähentämättömät sukusolut, jotka luokitellaan samoiksi lajeiksi.
Allopoliploidi on se, että organismi, joka sisältää ei -homologisten kromosomien pelejä, jotka johtuvat hybridisaatiosta eri lajien välillä. Tässä tapauksessa polyploidia tapahtuu kahden liittyvän lajin hybridisaation jälkeen.
Polyploidia eläimissä
Eläimissä polyploidia on harvinaista tai harvinaista. Yleisin levinnyt hypoteesi, joka selittää polyploidilajien pienen taajuuden suuremmilla eläimillä.
Tämä ajatus on ylläpidetty huolimatta kertyneistä todisteista eläimistä, jotka ovat polyploidina. Sitä havaitaan yleensä alemmissa eläinryhmissä, kuten matoissa ja monenlaisissa Plalmintosissa, joissa yksilöillä on yleensä sekä uros- että naisten sukurauhaset, jotka helpottavat itsekoristeista.
Lajeja, jotka esittävät tätä viimeistä tilaa. Toisaalta se voi esiintyä myös muissa ryhmissä, joiden naaraat voivat antaa jälkeläisiä ilman hedelmöitystä, partenogeneesiksi kutsutun prosessin kautta (joka ei tarkoita normaalia meioottista seksuaalista sykliä)
Parthenogenesin aikana jälkeläisiä tuotetaan periaatteessa vanhempien solujen mitoottinen jako. Täällä on monia selkärangattomia, kuten kovakuoriaisia, isopodeja, koita, katkarapuja, useita arachnidiryhmiä ja joitain kalalajeja, sammakkoeläimiä ja matelijoita.
Toisin kuin kasvit, eläimissä spesifikaatio polyploidian kautta muodostaa poikkeuksellisen tapahtuman.
Esimerkkejä eläimissä
Jyrsijä Tympanoctomys barrerae Se on tetraploidilaji, jolla on 102 kromosomia somaattista solua kohti. Siinä on myös "gigas" -vaikutus siittiöönsä. Tämä alopoliploidilaji on todennäköisesti peräisin muiden jyrsijälajien useiden hybridisaatiotapahtumien esiintymisestä Octomys Mimax ja Pipanacoctomys aureus.
Polyploidia ihmisillä
Polyploidia on harvinaista selkärankaisilla, ja sitä pidetään merkityksettömänä ryhmien, kuten nisäkkäiden (toisin kuin kasvien) monipuolistamisessa, sukupuolen määritysjärjestelmässä ja annoksen kompensointimekanismissa tapahtuvien häiriöiden vuoksi.
Arvioidaan, että viisi jokaisesta 1000 ihmisestä syntyy vakavia geneettisiä vikoja, jotka johtuvat kromosomaalisista poikkeavuuksista. Vielä suurempi määrä alkioita, joilla on kromosomaalisia vikoja, keskeytetään spontaanisti, ja monet eivät koskaan saavuta syntymää.
Voi palvella sinua: Transposonit: Mitkä ovat, tyypit, ominaisuudetIhmisillä kromosomaalisia polyploideja pidetään tappavina. Somaattisissa soluissa, kuten hepatosyyteissä, noin 50% näistä on normaalisti polyploidia (tetraploidia tai oktaploidia).
Lajien yleisimmin havaitut polyploidiat ovat täynnä triploidioita ja tetraploidioita, lisäksi sekoitettu diploidi/triploidi (2N/3N) ja diploidi/tetraploidi (2N/4N).
Jälkimmäisessä normaalien diploidisolujen (2N) populaatio on rinnakkain, toisella, jolla on 3 tai enemmän haploidikromosomeja, esimerkiksi: triploidi (3N) tai tetraploidi (4N).
Triploidia ja tetraplodioita ihmisillä eivät ole elinkelpoisia pitkällä aikavälillä. Useimmissa tapauksissa kuolemasta on ilmoitettu useimmissa tapauksissa tai jopa muutaman päivän kuluttua syntymästä, joka vaihtelee alle kuukaudesta enintään 26 kuukauteen.
Polyploidia kasveissa
Usean kuin yhden genomin olemassaololla samassa ytimessä on ollut tärkeä rooli kasvien alkuperässä ja evoluutiossa, ehkä tärkein sytogeneettinen muutos kasvien spesifikaatiossa ja evoluutiossa. Kasvit olivat sisäänkäynnin ovi solujen tietämykseen, joissa oli enemmän kuin kaksi kromosomipeliä solua kohti.
Kromosomimäärien alusta lähtien havaittiin, että monenlaisia villiä ja viljeltyjä kasveja (mukaan lukien jotkut tärkeimmistä) ovat polyploideja. Lähes puolet angiospermien lajeista (kasvit, joilla on kukilla) ovat polyploideja, samoin suurin osa saniaisista (95%) ja monenlaisia sammaleita.
Polyploidian esiintyminen kuntosalikasveissa on harvinaista ja erittäin vaihtelevaa angiosperms -ryhmissä. Yleensä on huomautettu, että polyploidikasvit ovat erittäin mukautuvia, kykenevät miehittämään elinympäristöjä, joita heidän diploidiset esivanhempansa eivät pystyneet. Lisäksi polyploidikasvit, joissa on enemmän genomisia kopioita, keräävät suuremman "variaatio".
Kasveissa ehkä alopoliploideilla (yleisempiä luonteeltaan) oli perustavanlaatuinen rooli monien ryhmien spesifikaatiossa ja adaptiivisessa säteilyssä.
Puutarhanhoito
Vihannesissa polyploidia voi olla peräisin useista erilaisista ilmiöistä, jotka ovat ehkä meioosiprosessin aikana yleisimpiä virheitä, jotka aiheuttavat diploidisia sukusoluja.
Yli 40% viljeltyistä kasveista on polyploideja, mukaan lukien sinimailasen, puuvillan, perunan, kahvin, mansikat, vehnä, ilman että kodistumisen ja kasvien polyploidian välillä on suhde.
Koska patja toteutettiin agenttina polyploidian indusoimiseksi, sitä käytettiin satokasveissa pohjimmiltaan kolmesta syystä:
-Polyploidian tuottamiseksi tietyissä tärkeyslajeissa, kuten yritys saada parempia kasveja, koska polyploideissa on yleensä fenotyyppi, jossa koon "gigas" on huomattava kasvu, koska soluja on suurempi määrä. Tämä on sallinut puutarhanhoito- ja kasvien geneettisen parantamisen alan merkittävän edistymisen.
Voi palvella sinua: Gene Flow-Hybridien polyploidisointiin ja että ne palauttavat hedelmällisyyden siten, että se on suunniteltu tai syntetisoitu mikä tahansa laji.
-Ja lopuksi keinona siirtää geenejä lajien välillä, joilla on erilainen ploidia, tai samassa lajissa.
Esimerkkejä kasveissa
Kasveissa luonnollinen polyploidi, jolla on suuri merkitys ja erityisen mielenkiintoinen, on leivän vehnä, Triticum aestibum (heksaploidi). Yhdessä rukiin, polyploidiksi, nimeltään "Triticale", allpoliploidi, jolla on korkea vehnän tuottavuus ja rukiin tukevuus, jolla on suuri potentiaali.
Vehnä viljeltyjen kasvien sisällä on ollut erittäin välttämätöntä. On 14 vehnälajia, jotka ovat kehittyneet alopolyloidialla, ja muodostavat kolme ryhmää, yksi 14: stä, toinen 28: sta ja yksi viimeinen 42 kromosomista. Ensimmäinen ryhmä sisältää suvun vanhimmat lajit T. Monococcum ja T. Bootticum.
Toisessa ryhmässä koostuu 7 lajista ja ilmeisesti johtuu hybridisaatiosta T. Bootticum Eräänlaisella villillä kiehumisella toisesta nimeltä genre Aegilops. Risteys tuottaa steriilin voimakkaan hybridi.
Kolmas 42 kromosomin ryhmä on siellä, missä leivän vehnät ovat, jotka todennäköisesti ovat peräisin hybridisoinnista sänkylajeista toisen lajin kanssa Aegilops jota seuraa kromosomaalinen komplementin päällekkäisyys.
Viitteet
- Alcántar, j. P. (2014). Polyploidia ja sen evoluutio -merkitys. Aiheuttaa puutetta ja tekniikkaa, 18: 17-29.
- Ballesta, f. J -. (2017). Joitakin bioeettisiä näkökohtia suhteessa ihmisten tapauksiin, joilla on täydellinen tetraploidia tai triploidia. Bioetica, 10(10): 67-75.
- Castro, S., & Loureiro, J. (2014). Lisääntymisen rooli polyploidikasvien alkuperässä ja kehityksessä. Ekosysteemilehti, 23(3), 67-77.
- Freeman, S ja Herron, J. C. (2002). Evoluutioanalyysi. Pearson -koulutus.
- Hichins, c. F. Yllyttää. (2010). Tetraploidisen jyrsijöiden tympanoctomys barrerae (OctoDontidae) geneettinen ja maantieteellinen alkuperä, joka perustuu mitokondriaalisten sytokondriaalisten sekvenssien analyysiin (Väitöskirjan tohtori, ekologian instituutti).
- Hickman, c. P, Roberts, L. S., S. Lens., Larson, a., I'anson, h. & Eisenhour, D. J -. (2008). Eläintieteen integroidut priormit. New York: McGraw-Hill. 14th Painos.
- Pimentel benítez, h., Lantigua curz,., & Quiñones Maza tai. (1999). Diploid-tetraploidi Mixoploidy: Ensimmäinen raportti ympäristössämme. Kuuban lastenlääketieteellinen aikakauslehti, 71(3), 168-173.
- Schifino-Wittmann, M. T. (2004). Polyploidia e Seu Impact Na Origin ja Evolução Das villi ja viljelty kasvit. Brasilian Agrociencian aikakauslehti, 10(2): 151-157.
- Suzuki, D. T.; Griffiths, a. J -. F.; Miller, J. H & Lewontin, R. C. (1992). Johdanto geenianalyysiin. McGraw-Hill-Amerikanvälinen. 4th Painos.
- « Euploidía alkuperä, tyypit ja seuraukset
- Chamelaucium Uncinatum -ominaisuudet, elinympäristö, hoito »