

Ydinhuokosominaisuudet, toiminnot, komponentit
- 599
- 147
- Dr. Travis Reichert
Hän ydinhuokos (Kreikasta, huokoset = askel tai kuljetus) on ytimen "ovi", joka sallii suurimman osan nukleoplasman ja sytoplasman välillä. Ydinhuokos liittyy ytimen sisäisiin ja ulkoisiin kalvoihin muodostaakseen kanavia, jotka palvelevat proteiinien ja RNA: n kuljettamista.
Sana poro ei heijasta kyseisen rakenteen monimutkaisuutta. Tämän vuoksi on parempi viitata ydinhuokoskompleksiin (CPN) ydinhuokosen sijasta. CPN voi kokea muutoksia rakenteessaan kuljetuksen aikana tai solusyklin tiloja.
Viime aikoina on havaittu, että CPN: n muodostavat nukleoporiinit, proteiinit, on tärkeä rooli geeniekspression säätelyssä. Siten, kun esiintyy mutaatioita, jotka vaikuttavat nukleoporiinien toimintaan, patologioita tuotetaan ihmisillä, kuten autoimmuuni, kardiomyopatiat, virusinfektiot ja syöpä.
[TOC]
Ominaisuudet
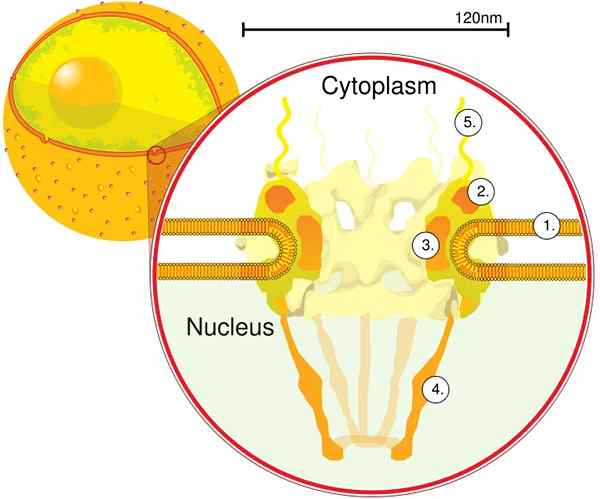
Elektronimomografian kautta määritettiin, että CPN: n paksuus on 50 nm, ulkoinen halkaisija välillä 80 - 120 nm ja sisähalkaisija 40 nm. Suuret molekyylit, kuten suuren ribosomin alayksikkö (PM 1590 kDa), voidaan viedä ytimen ulkopuolelle CPN: n läpi. On arvioitu, että ydintä on 2000 - 4000 CPN.
Yksittäisen CPN: n molekyylipaino on noin 120 - 125 MDA (1 MDA = 106 Da) selkärankaisilla. Sitä vastoin CPN on hiivassa pienempi, jossa siinä on noin 60 MDA. CPN: n valtavasta koosta huolimatta nukleoporiinit ovat erittäin säilyneet kaikissa eukaryooteissa.
Translokaalisointi CPN: n kautta on nopea prosessi, jonka nopeus on 1000 siirtoa/sekunti. CPN ei kuitenkaan määritä itse kuljetusvirtauksen suuntaa.
Tämä riippuu RangTP -gradientista, joka on ytimessä suurempi kuin sytosolissa. Tätä kaltevuutta ylläpitää guaniininvaihtimen tekijä.
Solusyklin aikana CPN: t kokevat kokoonpano- ja erimielisyyssyklit. Kokoonpano tapahtuu rajapinnassa ja heti mitoosin jälkeen.
Funktiot
Ribonukleiinihappo (pieni ydin -RNA, Messenger RNA, siirto -RNA), proteiinit ja ribonukleoproteiinit (RNP) on kuljetettava aktiivisesti CPN: n läpi. Tämä tarkoittaa, että ATP: n ja GTP -hydrolyysin energia vaaditaan. Jokainen molekyyli kuljetetaan tietyllä tavalla.
Yleensä RNA -molekyylit on täynnä proteiineja, jotka muodostavat RNP -kompleksit, jotka viedään tällä tavalla. Sitä vastoin ytimeen aktiivisesti kuljetettujen proteiinien on oltava sekvenssimerkki sijainnista ytimessä (SLN), aminohappojätteen hallussapitäjä positiivisella kuormalla (esimerkiksi KKKRK).
Voi palvella sinua: mesosomiYtimeen vietyjen proteiinien on oltava vientisignaali ytimeen (NES), joka on runsaasti leusiiniaminohapossa.
Ytimen ja sytoplasman välisen kuljetuksen helpottamisen lisäksi CPN: t osallistuvat kromatiinin organisointiin, geeniekspression säätelyyn ja DNA: n korjaukseen. Nukleoporiinit (NUPS) edistävät transkription aktivaatiota tai tukahduttamista solujen lisääntymistilasta riippumatta.
Hiivoissa nuptit löytyvät ydinkääreiden CNP: stä. Heidän sisällä olevissa metatsoarteissa. He suorittavat samat toiminnot kaikissa eukaryooteissa.
Aineiden tuonti
CPN: n kautta pienten molekyylien passiivinen leviäminen molemmissa suunnissa että aktiivisessa kuljetuksessa, proteiinin tuonnissa, RNA: n ja ribonukleoproteiinien (RNP) vienti ja kaksisuuntaiset molekyylit sukkula. Jälkimmäinen sisältää RNA: n, RNP: n ja proteiinien, jotka osallistuvat signalointiin, biogeneesiin ja korvaamiseen.
Proteiinien tuonti ytimessä tapahtuu kahdessa vaiheessa: 1) proteiini, joka sitoutuu CPN: n sytoplasmiseen puoleen; 2) ATP -riippuvainen translocalisaatio CPN: n kautta. Tämä prosessi tarvitsee ATP -hydrolyysin ja GTP/BKT: n vaihdon ytimen ja sytoplasman välillä.
Kuljetusmallin mukaan vastaanottava proteiinikompleksi liikkuu kanavaa pitkin unionin, dissosiaation ja liiton kautta nukleoporiinien toistuviin FG-sekvensseihin. Tällä tavoin kompleksi siirtyy yhdestä nukleoporiinista toiseen CPN: n sisällä.
Aineiden vienti
On samanlainen kuin tuonti. Ran GTPaasi asettaa suuntauksen kuljetukseen CNP: n kautta. RAN on molekyylikytkin, jossa on kaksi konformaatiotilaa, riippuen siitä, liittyykö se BKT: hen tai GTP: hen.
Kaksi spesifistä säätelyproteiinia laukaisee kahden tilan välisen konversion: 1) sytosolinen GTPaasi aktivoiva proteiini (GAP), joka tuottaa GTP-hydrolyysiä ja muuntaa siten RAN-GTP: n RAN-GDP: ksi; ja 2) Ydin Guanina -vaihtotekijä (GEF), joka edistää BKT: n vaihtoa GTP: llä ja muuntaa RAN-GKT: n RAN-GTP: ksi.
Sytosoli sisältää pääasiassa RAN-GKT: n. Ydin sisältää pääasiassa RAN-GTP: tä. Tämä RAN: n kahden konformaatiomuodon gradientti ohjaa kuljetusta asianmukaisessa osoitteessa.
Unioni helpottaa vastaanottimen tuontia yhdessä aseman kanssa toistoihin. Jos saavutat CNP: n ydinvoiman, RAN-GTP liittyy vastaanottimeen vapauttaaksesi sijaintisi. Siten RAN-GTP luo tuontiprosessin suunnan.
Ydinvienti on samanlainen. Ytimessä RAN-GTP edistää kuitenkin vientivastaanottajan aseman liiton. Kun vientivastaanotin liikkuu huokosen läpi kohti sytosolia, se täyttää RAN-raon, joka indusoi GTP-hydrolyysin BKT: hen. Lopuksi vastaanotin vapautuu hänen asemastaan ja juoksee-GKT: n sytosolissa.
Voi palvella sinua: polymorfonukleaariset leukosyytitRNA -kuljetus
Tietyntyyppisten RNA: n vienti on samanlainen kuin proteiinien vienti. Esimerkiksi ARNT ja RNANS (pieni ydin) käyttävät RangTP -gradienttia ja kuljetetaan CPN: n läpi keinoin. Kypsien ribosomien vienti on myös riippuvainen RangTP -gradientista.
MRNA viedään hyvin eri tavalla kuin proteiinit ja muut RNA: t. Vientiä varten mRNA muodostaa RNP Messenger (RNPM) -kompleksin, jossa RNM -molekyyliä ympäröivät sadat proteiinimolekyylit. Näillä proteiineilla on prosessointi, Enimmäismäärä, MNA: n Empalme ja polyadentilly.
Solun on kyettävä erottamaan RNPM kypsällä RNM: llä ja RNPM: llä epäkypsällä mRNA: lla. RPNM -kompleksin muodostava mRNA voisi hyväksyä topologiat, jotka on uudistettava kuljetukseen. Ennen kuin RNPM tulee CPN.
Kun kypsä RNPM kootaan, RPNM kuljetetaan kanavan kautta kuljetusvastaanottimen avulla (NXF1-NXT1). Tämä vastaanotin tarvitsee ATP -hydrolyysin (ei RANGTP -gradienttia) RNPM: n suuntauksen uudelleenmuodostumisen määrittämiseksi, joka saavuttaa sytoplasman.
Ydinhuokoskompleksi ja geeniekspression hallinta
Jotkut tutkimukset osoittavat, että CPN -komponentit voivat vaikuttaa geneettisen ekspression säätelyyn kontrolloimalla kromatiinirakennetta ja sen saavutettavuutta transkriptiotekijöihin.
Uuden evoluution eukaryooteissa heterokromatiini sijaitsee mieluummin ytimen reunalla. Tätä aluetta keskeyttää euchromatiinikanavat, joita ylläpitää CPN -ydinkori. Ydinkorin yhteys euchromatiiniin liittyy geenitranskriptioon.
Esimerkiksi transkription aktivointi CPN -tasolla merkitsee ydinkorin komponenttien vuorovaikutusta proteiineilla, kuten histoniasetyylitransferaasisaga- ja RNA -vientitekijät.
Siten ydinkori on alusta lukuisille huoltogeeneille (Taloudenhoito;.
Ydinhuokos- ja virologiakompleksi
Virus Eukaryot -soluinfektio riippuu CPN: stä. Kummassakin virusinfektion tapauksessa sen menestys riippuu CPN: n läpi kulkevasta DNA: sta, RNA: sta tai RPN: stä, jotta saadaan lopullinen tavoite, joka on viruksen replikaatio.
Voi palvella sinua: sytoplasmiset sulkeumat: mitkä ovat, ominaisuudet, toiminnotApe -virus 40 (SV40) on ollut yksi tutkituimmista malleista tutkiakseen CPN: n toimintaa translokaalisoinnissa ytimessä. Tämä johtuu siitä, että SV40: llä on pieni genomi (5.000 tukikohtaa).
On osoitettu, että virus -DNA: n kuljetus helpottaa viruksen proteiiniproteiineja, jotka suojaavat virusta, kunnes ydin saavuttaa.
Komponentit
CPN on upotettu ydinkuoreen ja koostuu välillä noin 500 - 1000 nuptia. Nämä proteiinit on järjestetty rakenteellisiin alakomplekseihin tai moduuleihin, jotka ovat vuorovaikutuksessa toistensa kanssa.
Ensimmäinen moduuli on keskuskomponentti tai rengas huokosen sisällä hiekkakellon muodossa, jota rajoittaa toinen halkaisijaltaan 120 nm: n rengas molemmin puolin, intranukleaarinen ja sytoplasminen. Toinen moduuli ovat ydin- ja sytoplasman renkaat (halkaisijaltaan 120 nm), joka sijaitsee Sandwate -muotoisen komponentin ympärillä.
Kolmas moduuli on kahdeksan filamenttia, jotka projisoidaan 120 nm: n renkaasta nukleoplasmassa ja muodostavat korin muotoisen rakenteen. Neljäs moduuli koostuu filamenteista, jotka projisoidaan sytoplasman sivulle.
Y -muotoinen kompleksi, joka koostuu kuudesta NUP: sta ja SEH 1: stä ja SEC 13 -proteiinista, on CNP: n suurin ja parhaiten karakterisoitu kompleksi. Tämä kompleksi on välttämätön yksikkö, joka on osa CPN -telineitä.
Huolimatta NUPS -sekvenssien alhaisesta samankaltaisuudesta, CPN -telineet säilyvät voimakkaasti kaikissa eukaryooteissa.
Viitteet
- Beck, m., E. 2016. Ydinhuokoskompleksi: sen toiminnan ymmärtäminen rakenteellisen näkemyksen avulla. Luontoarvostelut, molekyylisolubiologia, doi: 10.1038/NRM.2016.147.
- Ibarra, a., Hetzer, m.W -. 2015. Ydinhuokosproteiinit ja genomitoimintojen hallinta. Geenit ja kehitys, 29, 337-349.
- Kabachinski, G., Schwartz, t.TAI. 2015. Ydinhuokoskompleksi - rakenne ja toiminta yhdellä silmäyksellä. Journal of Cell Science, 128, 423-429.
- Knocknhauer, k.JA., Schwartz, t.TAI. 2016. Ydinhuokoskompleksi joustavana ja dynaamisena porttina. Solu, 164, 1162-1171.
- Ptak, c., Aitchison, J.D -d., Wozniak, r.W -. 2014. Monitoiminen ydinkompleksi: alusta geeniekspression hallitsemiseksi. Solubiologian nykyinen mielipide, doi: 10.1016/j.Ceb.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Vilkaisu: ydinhuokoskompleksi - kattava katsaus rakenteesta ja toiminnasta. International Journal of Academic Medicine, 3, 51-59.
- Tran, e.J -., S. S.R -. 2006. Kompleksien dynaaminen ydin: Elämä reunalla. Solu, 125, 1041-1053.