

Fylogeneettiset puutyypit ja ominaisuudet, esimerkkejä
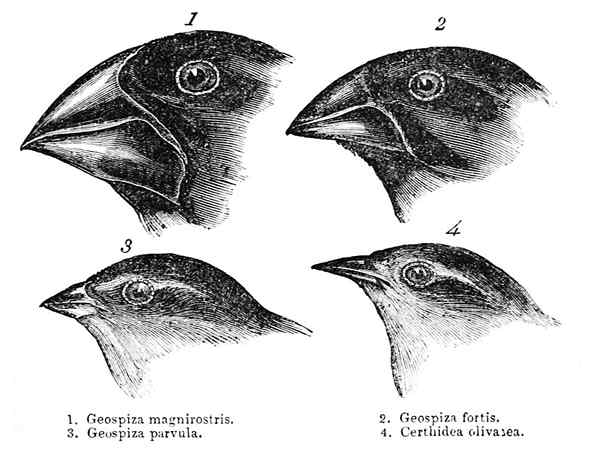
- 5159
- 67
- Edgar VonRueden
Eräs fylogeneettinen puu Se on historian ja esi -isän suhteiden matemaattinen graafinen esitys - ryhmien, populaatioiden, lajien tai minkä tahansa muun taksonomisen luokan jälkeläiset. Teoreettisesti kaikki fylogeneettiset puut voidaan ryhmitellä elämänpuuhun, muodostaen universaalin puun.
Nämä graafiset esitykset ovat mullistaneet evoluutiobiologian tutkimuksen, koska ne sallivat lajin perustamisen ja määrittelemisen, testaamalla erilaisia evoluutiohypoteeseja (kuten endosimbioottinen teoria), arvioimalla sairauksien (kuten HIV) alkuperää jne.
Lähde: John Gould (14.Sekoittaa.1804 - 3.Helmikuu.1881) [julkinen alue] Puut voidaan rakentaa uudelleen käyttämällä morfologisia tai molekyylihahmoja tai molemmat. Samoin on olemassa erilaisia menetelmiä niiden rakentamiseksi, yleisin on kladistien menetelmä. Tämän tarkoituksena on tunnistaa jaetut johdetut merkit, jotka tunnetaan nimellä Synapomorphia.
[TOC]
Ominaisuudet
Yksi Charles Darwinin kehittämistä periaatteista muodostaa kaikkien elävien organismien yhteisen esi -isän - ts. Meillä kaikilla on syrjäinen esi -isä.
Sisään "Lajien alkuperä" Darwin nostaa "elämänpuun" metaforin. Itse asiassa käytä hypoteettista graafista puuta ideasi kehittämiseen (uteliaana, se on ainoa esimerkki Alkuperä-A.
Tämän metaforin esitys on se, mitä tunnemme fylogeneettisiksi puiksi, joiden avulla voimme graafisesti osoittaa joidenkin betoniryhmien historiaa ja suhteita.
Fylogeneettisen puun anatomia
Fylogeneettisissä puissa voimme erottaa seuraavat osat - jatkaen kasvitieteellisellä analogialla:
Oksat: Puulinjoja kutsutaan "oksiksi", ja nämä edustavat tutkimuspopulaatioita ajoissa. Puun tyypistä riippuen (katso myöhemmin) oksan pituudella voi olla merkitys.
Haarojen kärjessä löydämme organismit, joita haluamme arvioida. Nämä voivat olla tällä hetkellä elossa olevia yhteisöjä tai sukupuuttoon kuolleita. Laji olisi puun lehdet.
Juuri: Juuri on puun esi -isien haara. Joillakin on se ja niillä kutsutaan juurtuneiksi puiksi, kun taas toisilla ei ole sitä.
Solmut: Haarojen haarapisteitä kahdessa tai useammassa linjassa kutsutaan solmuiksi. Kohta edustaa jälkeläisten ryhmien viimeisintä yleistä esi -isää (huomaa, että nämä esi -isät ovat hypoteettisia).
Solmun olemassaolo merkitsee spesifikaatiotapahtumaa - uusien lajien luomista. Tämän jälkeen jokainen laji seuraa evoluutiokurssia.
Ylimääräinen terminologia
Näiden kolmen peruskäsitteen lisäksi fylogeneettisten puiden suhteen on muita tarvittavia termejä:
Poliitio: Fylogeneettisessä puussa se esittelee enemmän kuin kaksi haaraa solmussa, sanotaan, että poliittinen. Näissä tapauksissa fylogeneettinen puu ei ole täysin ratkaistu, koska mukana olevien organismien väliset suhteet eivät ole selviä. Tämä tapahtuu yleensä tietojen puutteen vuoksi, ja se voidaan ratkaista vain, kun tutkija kertyy enemmän.
Voi palvella sinua: tetrosas: ominaisuudet, erythrosa, synteesi, johdannaisetUlkoinen ryhmä: Fylogeneettisissä teemoissa on yleistä kuunnella ulkoisen ryhmän käsitettä - jota kutsutaan myös Ryhmäryhmä. Tämä ryhmä on valittu pystyvän juurtumaan puuhun. Se on valittava taksoniksi, joka aikaisemmin poikkesi tutkimusryhmää. Esimerkiksi, jos opiskelen Echinodermoa, voit käyttää Ryhmäryhmä ascidias.
Kaverit
Puita on kolme perustyyppiä: kladogrammit, lisäaineet ja ultrapohjaiset puut.
Clayogrammit ovat yksinkertaisimpia puita ja osoittavat organismien suhdetta yhteisessä esi -isissä. Tämän tyyppisen puun tiedot ovat haarautumismalleissa, koska oksien kolla ei ole ylimääräistä merkitystä.
Toinen puutyyppi on lisäaine, jota kutsutaan myös metrisiksi puiksi tai philogrammiksi. Oksojen pituus liittyy evoluutiomuutoksen määrään.
Lopuksi, meillä on ultrametrisiä puita tai dendogrammeja, joissa puiden kaikki kärjet ovat samalla etäisyydellä (mitä ei tapahdu filogrammissa, missä kärki voi näkyä alapuolella tai korkeampi kuin kumppanisi). Haaran pituus liittyy evoluutioaikaan.
Puun valinta liittyy suoraan evoluutiokysymykseen, johon haluamme vastata. Esimerkiksi, jos välitämme vain yksilöiden välisistä suhteista, kladogrammi riittää tutkimukseen.
Yleisimmät virheet fylogeneettiset puita
Vaikka fylogeneettiset puut ovat yleensä grafiikkaa laaja -alaisesta evoluutiobiologiassa (ja yleensä biologiassa), on monia opiskelijoita ja ammattilaisia, jotka ymmärtävät väärin viestin, että nämä grafiikat - ilmeisesti yksinkertaiset - aikovat toimittaa lukijalle.
Tavaratilaa ei ole
Ensimmäinen virhe on lukea ne sivusuunnassa, olettaen, että evoluutio merkitsee etenemistä. Jos ymmärrämme evoluutioprosessin oikein, ei ole syytä ajatella, että vasemmalla ovat esi -isien ja oikean lajin edistyneimmät lajit.
Vaikka puun kasvitieteellinen analogia on erittäin hyödyllinen, on piste, jossa se ei ole enää niin tarkka. Puussa on ratkaiseva rakenne, jota ei ole puussa: tavaratila. Fylogeneettisissä puissa emme löydä päähaaraa.
Erityisesti jotkut ihmiset voisivat pitää ihmistä evoluution "perimmäisenä tavoitteena", ja siksi lajit Homo sapiens Sen tulisi aina sijaita lopullisena kokonaisuutena.
Voi palvella sinua: Kasvaimen nekroositekijä (TNF): rakenne, toimintamekanismi, toimintaTämä visio ei kuitenkaan ole yhdenmukainen evoluutioperiaatteiden kanssa. Jos ymmärrämme, että fylogeneettiset puut ovat liikkuvia elementtejä, voimme sijoittaa Homo Missä tahansa puun terminaalissa, koska tämä ominaisuus ei ole merkityksellinen esityksessä.
Solmut voivat pyöriä
Tärkeä ominaisuus, joka meidän on ymmärrettävä fylogeneettisistä puista, on, että ne edustavat ei -staattista grafiikkaa.
Niissä kaikki nämä oksat voivat pyöriä - samalla tavalla kuin matkapuhelin voi tehdä sen. Emme tarkoita, että oksat voidaan siirtää alkuperäiskansoihimme, koska jotkut liikkeet tarkoittaisivat kuvion muutosta tai Topologia Puusta. Mitä voimme kiertää, ovat solmut.
Puun viestin tulkitsemiseksi emme saa keskittyä oksien kärkeen, meidän on tehtävä se haarapisteissä, jotka ovat graafisen tärkein osa.
Lisäksi meidän on otettava huomioon, että puun piirtämiseen on olemassa monia tapoja. Monta kertaa se riippuu kirjan tai lehden tyylistä ja oksien muodon ja sijainnin muutokset eivät vaikuta tietoihin, joita he haluavat välittää meille.
Emme voi päätellä esi -isien tai "vanhojen" lajien olemassaoloa
Kun aiomme viitata lajeihin nykyinen Emme saa soveltaa esi -isien konnotaatioita. Esimerkiksi, kun ajattelemme simpanssien ja ihmisten välisiä suhteita, voisimme ymmärtää virheellisesti, että simpanssit ovat esi -isiä suvun suhteen suhteen.
Simpanssien ja ihmisten yhteinen esi -isä ei kuitenkaan ollut kumpikaan niistä. Ajatella, että simpanssi on esi -isä, oletetaan, että sen kehitys pysähtyi, kun molemmat sukulaiset on erotettu.
Näiden ideoiden saman logiikan jälkeen fylogeneettinen puu ei tarkoita, onko nuoria lajeja. Koska alleeliset taajuudet ovat jatkuvasti ja ajan myötä muuttuu uusia merkkejä, lajin ikä on vaikea määrittää, ja varmasti puu ei anna meille tällaisia tietoja.
"Alleelitaajuuksien muutos ajan myötä" on tapa, jolla populaatioiden genetiikka määrittelee evoluution.
Ne ovat muuttumattomia
Kun tarkkailemme fylogeneettistä puuta. Jos lisäämme lisää merkkejä puuhun, se muuttaa sen topologiaa.
Tutkijoiden asiantuntemus valittaessa parhaat hahmot, jotka sallivat selvittää kyseisten organismien suhteet ovat avainasemassa. Lisäksi on erittäin tehokkaita tilastollisia työkaluja, joiden avulla tutkijat voivat arvioida puita ja valita kaikkein uskottavia.
Voi palvella sinua: proliini: ominaisuudet, rakenne, toiminnot, ruokaEsimerkit
Kolme elämän aluetta: Archaea, bakteerit ja Eukarya
Vuonna 1977 tutkija Carl Woese ehdotti elävien organismien ryhmälle kolmella alueella: Archaea, Bakteerit ja Eukarya. Tämä uusi luokittelujärjestelmä (aikaisemmin oli vain kaksi luokkaa, Eukaryota ja Prokaryota) perustui ribosomaaliseen kylkiluun molekyylimarkkeriin.
Bakteerit ja eukaryootit ovat laajalti tunnettuja organismeja. Arkaat ovat yleensä sekoitettu bakteereihin. Ne eroavat kuitenkin syvästi solukomponenttiensa rakenteesta.
Siksi, vaikka ne ovat mikroskooppisia organismeja, kuten bakteereja, Archaea -domeenin jäsenet liittyvät enemmän eukaryooteihin - koska heillä on esi -isä lähempänä yhteistä.
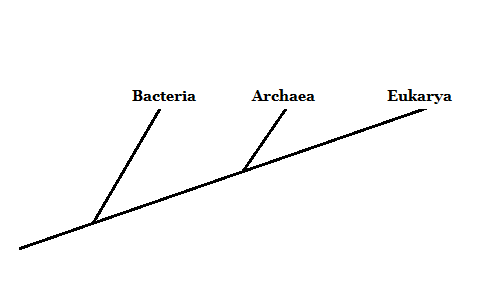 Lähde: Valmistaja Mariana Gelambi.
Lähde: Valmistaja Mariana Gelambi. Kädellisten filogeneesi
Evoluutiobiologian sisällä yksi kiistanalaisimmista kysymyksistä on ihmisen kehitys. Tämän teorian vastustajille ei ole logiikkaa evoluutiolle, joka perustuu Simiescon esi -isään, joka antoi nykyisen miehen.
Tärkein käsite on ymmärtää, että emme kehitty nykyisistä apinoista, vaan että meillä on yhteinen esi -isä heidän kanssaan. Apinoiden ja ihmisten puussa se korostaa, että se, mitä tunnemme "apinaksi", ei ole pätevä monofiilettinen ryhmä, koska se sulkee pois ihmisen.
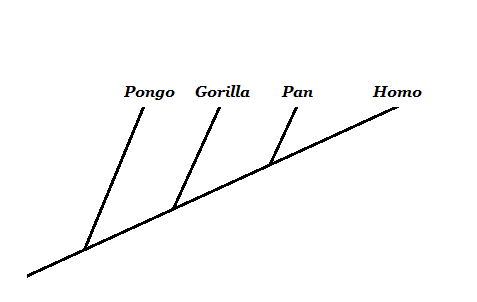 Lähde: Valmistaja Mariana Gelambi.
Lähde: Valmistaja Mariana Gelambi. CeTatorDACTILOS: n fylogeneesi (cetarioDACTYLA)
Evolutiongessa sanottu. Morfologisesti valaissa, delfiineillä ja muilla jäsenillä on vähän yhtäläisyyksiä muiden nisäkkäiden kanssa.
Tällä hetkellä erilaisten morfologisten ja molekyylisten luonteen tutkimuksen ansiosta.
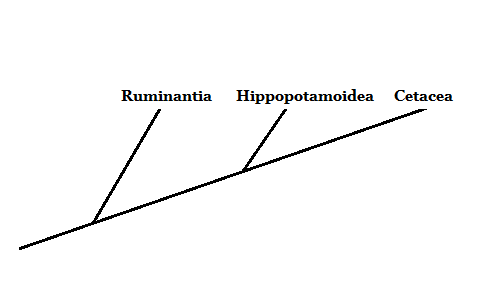 Lähde: Valmistaja Mariana Gelambi.
Lähde: Valmistaja Mariana Gelambi. Viitteet
- Baum, D. -Lla., Smith, S. D -d., & Donovan, S. S. (2005). Puuta ajatteleva haaste. Tiede, 310(5750), 979-980.
- Curtis, H., & Barnes, N. S. (1994). Kutsu biologiaan. Macmillan.
- BILL, B. K -k -. (Ed.-A. (2012). Homologia: vertailevan biologian hierarkkinen perusta. Akateeminen lehdistö.
- Hickman, c. P., Roberts, L. S., Larson, a., Ober, w. C., & Garrison, c. (2001). Eläintieteen integroidut priormit. McGraw-Hill.
- Kardong, k. V. (2006). Selkärankaiset: vertaileva anatomia, funktio, evoluutio. McGraw-Hill.
- Kliman, r. M. (2016). Evoluutiobiologian tietosanakirja. Akateeminen lehdistö.
- Losos, j. B -. (2013). Princeton -opas evoluutioon. Princeton University Press.
- Sivu, r. D -d., & Holmes ja. C. (2009). Molekyylin evoluutio: fylogeneettinen lähestymistapa. John Wiley & Sons.
- Riisi, s. -Lla. (2009). Evoluution tietosanakirja. Tiedottaa julkaisemisesta.
- Starr, c., Evers, c., & Starr, L. (2010). Biologia: Käsitteet ja sovellukset ilman fysiologiaa. Cengage -oppiminen.