

Lipidityyppien synteesi ja niiden päämekanismit
- 4287
- 162
- Gustavo Runte DVM
Se Lipidisynteesi Se koostuu sarjasta entsymaattisia reaktioita, joiden avulla lyhyen ketjun hiilivedyt tiivistetään muodostamaan pidempiä ketjumolekyylejä, jotka voivat myöhemmin kärsiä erilaisista kemiallisista modifikaatioista.
Lipidit ovat hyvin monipuolisia biomolekyylejä, jotka kaikki elävät solut syntetisoivat ja ovat erikoistuneet useisiin välttämättömiin toimintoihin solujen ylläpitämiseksi.
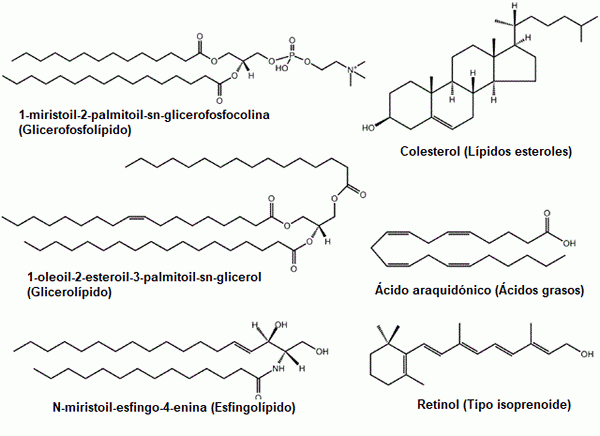 Joitakin esimerkkejä yleisistä lipideistä: glyserofosfolipidit, sterolit, glyserolipidit, rasvahapot, sfingolipidit ja prenolit (lähde: alkuperäinen lataus oli lmaps englanniksi wikipedia. / GFDL 1.2 (http: // www.GNU.Org/lisenssit/vanhojen lisenssit/FDL-1.2.HTML) Commonsin kautta, mukauttama Raquel Parada)
Joitakin esimerkkejä yleisistä lipideistä: glyserofosfolipidit, sterolit, glyserolipidit, rasvahapot, sfingolipidit ja prenolit (lähde: alkuperäinen lataus oli lmaps englanniksi wikipedia. / GFDL 1.2 (http: // www.GNU.Org/lisenssit/vanhojen lisenssit/FDL-1.2.HTML) Commonsin kautta, mukauttama Raquel Parada) Lipidit ovat biologisten kalvojen pääkomponentteja, mikä tekee niistä perustavanlaatuisia molekyylejä solujen olemassaoloon heidän ympäristönsä eristetyinä kokonaisuuksina.
Joillakin lipideillä on myös erikoistuneita toimintoja, kuten pigmenttejä, kofaktoreita, kuljettajia, pesuaineita, hormoneja, ekstracellulaarisia lähettiläitä, kovalenttisia ankkureita kalvoproteiineille jne. Siksi kyky syntetisoida erityyppisiä lipidejä on kriittinen kaikkien elävien organismien selviytymiselle.
Tämä suuri yhdisteryhmä luokitellaan perinteisesti useisiin luokkiin tai alaryhmiin: rasvahapot (kylläiset ja tyydyttymättömät), glyseridikompleksit lipidit (lipoproteiinit).
[TOC]
Lipidityypit ja niiden tärkeimmät synteesimekanismit
Kaikki lipidien biosynteesireitit ovat endogonisia ja pelkistäviä. Toisin sanoen, he kaikki käyttävät ATP: tä energian lähteenä ja vähentyneenä elektronin kuljettajana, kuten NADPH, pelkistävänä tehona.
Seuraavaksi kuvataan päälipidityyppien biosynteettisten reittien pääreaktiot, toisin sanoen rasva- ja eikosanoidihappo, triasyyliglyserolit ja fosfolipidit ja sterolit (kolesteroli) (kolesteroli) (kolesteroli) (kolesteroli) (kolesteroli).
- Rasvahapposynteesi
Rasvahapot ovat erittäin tärkeitä molekyylejä lipidien näkökulmasta, koska ne ovat osa solujen merkityksellisimpiä lipidejä. Hänen synteesi, toisin kuin monet tutkijat ajattelivat ensimmäisen tutkimuksen aikana, ei koostu heidän β-hapettumisensa käänteisestä reitistä.
Itse asiassa tämä metabolinen reitti esiintyy eri soluosastoissa ja vaatii kolmen hiiliatomin välittäjää, joka tunnetaan nimellä malonyyli-CoA, mikä ei ole välttämätöntä hapettumisessa.
 Malonyyli-CoA. Neurooteker / pub -alue
Malonyyli-CoA. Neurooteker / pub -alue Lisäksi se liittyy läheisesti asyyliryhmän kuljettajina tunnetuihin sulfhydriiliryhmiin (ACP, englanti Asyyli kantajaproteiinit-A.
Yleensä rasvahappojen, etenkin pitkän ketjun, synteesi on peräkkäinen prosessi, jossa jokaisessa ”paluu” toistetaan neljä vaihetta, ja kunkin paluun aikana on tyydyttynyt happaryhmä, joka on seuraavalle substraatti, joka on seuraavalle tarkoittaa uutta kondensoitumista uudella malonyyli-CoA-molekyylillä.
Jokaisessa vuoro- tai reaktiosyklissä rasvahappoketju ulottuu kahta hiilyä, kunnes pituus 16 atomia (palmitaatti) saavutetaan, minkä jälkeen sykli lähtee.
Malonyyli-CoA-muodostuminen
Tämä kolmen hiiliatomien välittäjä muodostuu peruuttamattomasti asetyyli-CoA: sta asetyyli-CoA-karboksylaasientsyymin vaikutuksen ansiosta, jolla on biotiinin eturauhasryhmä, joka on kovalenttisesti yhteydessä entsyymiin ja joka osallistuu tässä katalyysissä kahdessa vaiheessa kahdessa vaiheessa.
Tässä reaktiossa bikarbonaattimolekyylistä (HCO3-) johdettu karboksyyliryhmä siirretään biotiiniin ATP-riippuvaisessa muodossa, jossa biotinyyliryhmä täyttää molekyylin "väliaikaisen kuljetin" toiminnan siirrettynä acetyyli-CoA: lle, Malonyyli-CoA: n tuottaminen.
Voi palvella sinua: Oksidaasitesti: Perusta, menettely ja käyttöRasvahappojen synteesisekvenssissä käytetty pelkistävä aine on NADPH ja aktivoivat ryhmät ovat kaksi tioliryhmää (-sh), jotka ovat osa monientsimaattista kompleksia, jota kutsu.
Selkärankaisilla syntaasirasvahappokompleksi on osa yhtä suurta polypeptidiketjua, jossa esitetään 7 synteesireitille ominaisia entsymaattista aktiivisuutta, samoin kuin hydrolyyttistä aktiivisuutta, joka tarvitaan välittäjien vapauttamiseen synteesin lopussa.
 Syntasa-rasvahappoentsyymirakenne (lähde: Boehringer Ingelheim/CC BY-SA (https: // creativecommons.Org/lisenssit/by-SA/4.0) Wikimedia Commonsin kautta)
Syntasa-rasvahappoentsyymirakenne (lähde: Boehringer Ingelheim/CC BY-SA (https: // creativecommons.Org/lisenssit/by-SA/4.0) Wikimedia Commonsin kautta) Tämän kompleksin 7 entsymaattista aktiivisuutta ovat: ACILO-ryhmän kuljetusproteiini (ACP), asetyyli-CoAA-ACP transasetilas cetoasiili-ACP-reduktaasi (KR), β-hydroksasiili-ACP-dehydratasa (HD) ja vihan ACP-reduktaasi (ER).
Ennen kuin kondensaatioreaktiot voivat tapahtua rasvahappoketjun koottamiseksi, entsymaattisen kompleksin kaksi tioliryhmää ”ladataan” asyyliryhmien kanssa: Ensinnäkin asetyyli -COA siirretään kysteiinin -sh -ryhmään Kompleksin β-kettoasyyli-ACP-syntaasissa reaktio katalysoi asetyyli-CoAA-ACP-transsakietyentsyymi (AT) (AT).
Myöhemmin maloninen ryhmä siirretään malonyyli -CoA -molekyylistä ACILO -ryhmän (ACP) -kuljettimen -sh -ryhmään on osa rasvahappokompleksin syntaasia.
Neljän reaktion sekvenssi jokaiselle reaktiosyklin "palautukselle" on seuraava:
- Kondensaatio: Asetyyli- ja malonyyliryhmät, jotka "ladataan" entsyymissä, tiivistetään asetoasetyyli -ACP -molekyylin muodostamiseksi, joka on kytketty ACP -osaan ryhmän kautta -sh: n kautta. Tässä vaiheessa on CO2.
- Karbonyyliryhmän pelkistys: Karbonyyliryhmä asetoasetyyli-ACP: n C3-asennossa pelkistetään muotoon D-β-hydroksibutiril-ACP, reaktio, jota katalysoi β-pethoacyyli-ACP-reduktaasi, jota Nadph käyttää elektronin luovuttajana.
- Dehydraatio: D-β-hydroksibutiril-ACP: n C2- ja C3-hiilihiilet eivät ole vesimolekyylejä, muodostaen kaksoissidoksen, joka päättyy uuden yhdisteen tuotantoon trans-∆2-Butenoil-ACP. Tätä prosessia välittää β-hydroksiasyyli-ACP-dehydraasi (HD) -entsyymi (HD).
- Kaksinkertainen sidoksen pelkistys: Dehydraatiovaiheessa muodostetun yhdisteen kaksoissidos on tyydyttynyt (pelkistetty) saadaan aikaan Butiril-ACP: tä reaktiolla, jota katalysoi entsyymi Anger-ACP-reduktaasi (ER), joka käyttää myös NADPH: ta pelkistävänä aineena.
Synteesireaktioita esiintyy, kunnes muodostuu palmitaattimolekyyli (16 hiiliatomia), joka hydrolysoituu entsymaattisesta kompleksista ja joka vapautetaan mahdollisena edeltäjänä rasvahapoissa, joilla endoplasmisen retikulumin osa ja mitokondrioissa.
Se voi palvella sinua: Perun meren eläimistö ja kasvistoMuut modifikaatiot, joihin nämä molekyylit voivat kärsiä.
- Eikosanoidien synteesi
Eikosanoidit ovat solujen lipidejä, joilla on toimintoja, kuten ”lyhyen alueen” lähettilomolekyylit, jotka joidenkin kankaiden tuottama kommunikointi naapurimaiden kankaidensa kanssa. Nämä molekyylit syntetisoidaan 20 hiiliatomin monityydyttymättömistä rasvahapoista.
Prostaglandiinit
Vasteena hormonaaliseen ärsykkeeseen fosfolipaasientsyymi hyökkää kalvofosfolipideihin ja vapauttaa Araquidonato Glyserol Carbon 2: sta. Tämä yhdiste muunnetaan prostaglandiineiksi sileän endoplasmisen retikulumin entsyymin ansiosta, jolla on bifunktionaalinen aktiivisuus: syklooksigenaasi (COX) tai prostaglandiini H2 -syntaasi.
Tromboksaani
Prostaglandiinit voidaan muuntaa tromboksalaisiksi verihiutaleissa syntyneen tromboksaanin ansiosta (trombosyytit). Nämä molekyylit osallistuvat veren hyytymisen alkuvaiheisiin.
- Triakyyliglyserolien synteesi
Rasvahapot ovat perustavanlaatuisia molekyylejä muiden solujen monimutkaisempien yhdisteiden synteesiä varten, kuten triasyyliglyserolit tai lipidiglyserofosfolipidilipidikalvo (prosessit, jotka riippuvat solujen metabolisista tarpeista).
Eläimet tuottavat kolmesta yleisestä esiasteen triasyyliglyseroleista ja glycefosfalipideista: Fat-CoA-asyyli ja 3-fosfaatti L-glyseroli. Rasva-CoA-asyyli tuottaa asyyli-CoA-synteesat, jotka osallistuvat β-hapettumiseen, kun taas 3-fosfaatti L-glyseroli saadaan glykolyysistä ja kahden vaihtoehtoisen entsyymin vaikutuksesta: 3-fosfaattiglyserolidehydrogenaasi- ja kinaasiglyseroli.
Triakyyliglyserolit muodostuvat reaktiolla kahden Flo-CoA ACIL: n molekyylin ja 3-fosfaattiakyyliglyserolimolekyylin välillä; Nämä siirtoreaktiot katalysoivat spesifiset transferasit.
Tässä reaktiossa on alun perin fosfatidihappo, joka fosforyloituu entsyymifosfatidihappofosfataasilla tuottamaan 1,2-diayyliglyserolia, joka pystyy jälleen hyväksymään kolmannen rasva-CoA-asyylin molekyylin tuottavan triacilglyseroll.
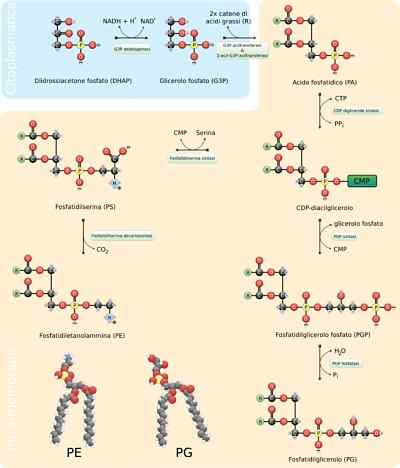
- Fosfolipidien synteesi
Fosfolipidit ovat erittäin vaihtelevia molekyylejä, kuten monet erilaiset voivat muodostua rasvahappojen ja "pää" -ryhmien yhdistelmällä glyseroliruenton (glysofosfolipidien) tai sfinosiinin (sfingolipidien) kanssa, jotka karakterisoivat niitä.
Näiden molekyylien yleiskokoonpano vaatii glyserolin tai pallomaisen luurankojen synteesin vastaavien rasvahappojen kanssa joko sterioinnilla tai amidoinnilla, lisäämällä hydrofiilisen ”pää” -ryhmän fosfodiéster -sidoksen läpi ja tarvittaessa, tarvittaessa , näiden viimeisten ryhmien muutos tai vaihto.
Eukaryooteissa tämä prosessi tapahtuu sileässä endoplasmisessa retikulumissa ja myös sisäisessä mitokondriaalisessa kalvossa, missä ne voivat pysyä määrittelemättömästi tai mistä ne voidaan siirtää muihin paikkoihin.
Reaktiovaiheet
Glyserofosfolipidien synteesireaktion ensimmäiset vaiheet ovat vastaavat triasyyliglyserolien tuotannon reaktioiksi, koska 3-fosfaattiglyserolimolekyyli esteröidaan kahteen rasvahappomolekyyliin hiilissä 1 ja 2 muodostaen fosfatidihappohapon. On yleistä löytää fosfolipidejä, joilla on tyydyttyneitä rasvahappoja C1: ssä ja jotka on tyydyttymättömät glyserolin C2: ssa.
Voi palvella sinua: ResistinaFosfatidihappo voidaan tuottaa myös diasyyliglyserolimolekyylin fosforylaatiolla, joka on jo syntetisoitu tai "kierrätetty".
Näiden molekyylien "pää" polaariryhmät muodostetaan fosfodiéster -linkkien kautta. Ensimmäinen asia, jonka pitäisi tapahtua tämän prosessin olevan oikein.
Jos tämä molekyyli sitoutuu diasyyliglyseroliin, niin muodostuu CDP-diakyyliglyseroli (fosfatidihapon "aktivoitu" muoto), mutta tämä voi tapahtua myös ryhmän "Head" -hydroksyyliryhmässä "Head".
Esimerkiksi fosfatidyyliseriinin tapauksessa diasilglyseroli aktivoidaan fosfatidihapon molekyylin kondensaatiolla trif-diakyylglyserolin muodostaen trif-sitidiinisyidiinimolekyylin (CTP) kanssa, muodostaen pyrofosfaatin ja eliminoimalla pyrofosfaatin.
Jos MMP-molekyyli (syytidiinimonofosfaatti) liikkuu seriinin tai hydroksyylihydroksyylin nukleofiilisen hyökkäyksen läpi 3-fosfaattiglyserolin hiili 1.
Molemmat tällä tavalla tuotetut molekyylit toimivat esiasteina muille kalvolipideille, joilla on usein biosynteettisiä reittejä toisistaan.
- Kolesterolisynteesi
Kolesteroli on olennainen molekyyli eläimille, jotka sen solut voivat syntetisoida, joten se ei ole välttämätöntä päivittäisessä ruokavaliossa. Tämä 27 hiiliatomin molekyyli tuotetaan edeltäjästä: asetaatti.
Tämä kompleksi molekyyli muodostuu asetyyli-CoA: sta neljässä päävaiheessa:
- Kolmen asetaattiyksikön kondensaatio muodostaa mevalonato, 6 hiilihytkien välittäjämolekyyli (ensin asetyyli-asetyyli-CoA-molekyyli muodostuu kahdella asetyyli-CoA: lla (tiolaasientsyymi) ja sitten toisella β-hydroksi-β-metyyliglutaril-CoA: sta (HMG-HYDroksi-β-metyyli COA) (HMG-CoA Synteteta -entsyymi). Mevalonato muodostuu HMG-CoA: sta ja HMG-CoA-reduktaasin entsyymin ansiosta.
- Mevalonato -muuntaminen isopreeniyksiköiksi. Ensimmäiset 3 fosfaattiryhmää siirretään 3 ATP -molekyylistä Mevalonatoon. Yksi fosfaateista menetetään yhdessä viereisen karbonyyliryhmän kanssa ja muodostetaan
- 6 hiiliatomin 6 isopreeniyksikön polymerointi tai kondensaatio 30 -hiiliatomien (lineaarinen molekyyli) parranajo (lineaarinen molekyyli) muodostamiseksi (lineaarinen molekyyli) muodostamiseksi.
- Escualean -sikraatio muodostaa kolesterolin steroidisen ytimen 4 renkaat ja sitä seuraavat kemialliset muutokset: metyyliryhmien hapetukset, migraatio ja eliminointi jne., Mikä tuottaa kolesterolia.
Viitteet
- Garrett, r. H., & Grisham, c. M. (2001). Biokemian periaatteet: ihmisen keskittymällä. Brooks/Cole Publishing Company.
- Murray, r. K -k -., Granner, D. K -k -., Mayes, P. -Lla., & Rodwell, V. W -. (2014). Harperin kuvitettu biokemia. McGraw-Hill.
- Nelson, D. Lens., Lehninger, a. Lens., & Cox, M. M. (2008). Lehninger -biokemian periaatteet. Macmillan.
- Jacquemyn, J., Cascaho, a., & Goodchild, r. JA. (2017). Endoplasmisen retikulumin kontrolloidun lipidi -biosynteesin endoplasmisen retikulumin edut ja out. EMBO Reports, 18 (11), 1905-1921.
- Ohlrogge, J., & Selaa, J. (tuhatyhdeksänsataayhdeksänkymmentäviisi). Lipidi -biosynteesi. Kasvisolu, 7 (7), 957.
- « José López Portillon elämäkerta, hallitus ja maksut
- Trooppinen viidakko Meksikon ominaisuuksissa, sijainti, helpotus, kasvisto, eläimistö »