

DNA -prosessin translaatio eukaryotasissa ja prokaryooteissa
- 3517
- 959
- Arthur Terry II
Se DNA -käännös Se on prosessi, jolla transkription aikana tuotettujen lähettiläiden tiedot.
Solujen näkökulmasta geenin ilmentyminen on suhteellisen monimutkainen aine, joka tapahtuu kahdessa vaiheessa: transkriptio ja translaatio.
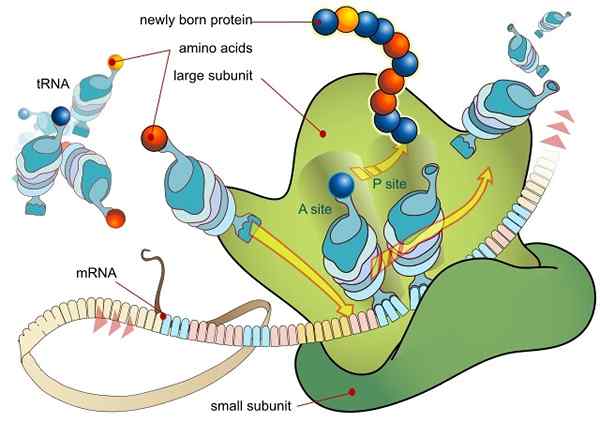
 RNA -translaatio, jota välittää ribosomi (lähde: ladyofhats / julkinen alue, Wikimedia Commonsin kautta)
RNA -translaatio, jota välittää ribosomi (lähde: ladyofhats / julkinen alue, Wikimedia Commonsin kautta) Kaikki ekspressoituja geenejä (riippumatta siitä, onko peptidisekvenssien, ts. Proteiinien koodaava vai ei), tekevät niin siirtämällä sen DNA -sekvenssiin sisältyvät tiedot Messenger Mer -molekyyliin (RNAM) prosessin kautta, jota kutsutaan transkriptioksi, nimeltään transkriptio.
Transkriptio saavutetaan erityisillä entsyymeillä, jotka tunnetaan nimellä RNA-polymeraasit, jotka käyttävät yhtä geenin DNA: n komplementaarisia juosteita muotina ”pre-arnm” -molekyylin synteesiä varten, joka myöhemmin syytetään kypsän kypsän kypsän muodostamiseksi muodostamaan kypsän kypsän kypsään.
Proteiineja koodaaville geeneille kypsän RNM: n sisältämät tiedot ”luetaan” ja käännetään aminohapoissa geneettisen koodin mukaisesti, joka määrittelee, mikä kodoni- tai nukleotidikolmio vastaa kutakin tiettyä aminohappoa.
Proteiinin aminohapposekvenssin spesifikaatio riippuu siis DNA: n typpe -emäksen alkuperäisestä sekvenssistä, joka vastaa geeniä ja sitten RNA: ssa, joka kuljettaa mainittua tietoa ytimestä sytosoliin (eukaryoottisoluissa); prosessi, joka määritellään myös RNM: n ohjaaman proteiinin synteesiksi.
Ottaen huomioon, että DNA: n ja RNA: n ja vain 20 aminohappoa muodostavat 4 typpialusta on 64 mahdollista yhdistelmää, eri kolmiot (kodonit) voidaan koodata aminohappo, joten sanotaan, että geneettinen koodi on " rappeutunut "(lukuun ottamatta aminohapposionionia, jonka koodaa ainutlaatuinen AUG -kodonia).
[TOC]
Eukaryota-käännös (vaiheet-prosessit)
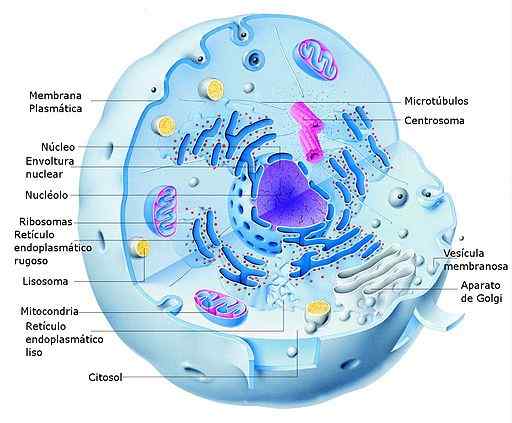
Eläinten eukaryot -solun ja sen osien kaavio (lähde: Alejandro Porto [CC0] Wikimedia Commonsin kautta) Eukaryoottisissa soluissa transkriptio tapahtuu ytimessä ja translaatio sytosoliin siten, että ensimmäisen prosessin aikana muodostuneet RNM: t täyttävät myös funktion tiedon kuljetuksessa ytimestä sytosoliin, jossa biosynteesikoneet (ribosomit -A.
On tärkeätä mainita, että transkription ja translaation osastojen jakaminen eukaryooteissa on totta ytimelle, mutta se ei ole sama organeleille, joilla on omat genominsa, kuten kloroplastit ja mitokondriot, joilla on enemmän samanlaisia järjestelmiä kuin prokaryoottisten organismejen järjestelmät kuin prokaryoottisten organismien kanssa kuin prokaryoottiset organismit.
Eukaryoottisoluissa on myös sytosolisia ribosomeja, jotka on kiinnitetty endoplasmisen retikulumin (karkea endoplasminen retikulum) kalvoihin, joissa proteiinien translaatio, joka on tarkoitettu asetettuna solumembraaneihin tai jotka vaativat translaation jälkeistä prosessointia.
- RNM -prosessointi ennen niiden käännöstä
RNM: ää muokataan niiden päissä, kun ne kirjoitetaan:
- Kun RNM: n 5'-pää syntyy RNA-polymeraasi II: n pinnalta transkription aikana, tätä "hyökkää" välittömästi entsyymien ryhmä, joka syntetisoi 7-metyyliguaniladon "konepellin" ja joka on kytketty terminaaliin RNM: n nukleotidi 5 'triffosfaattilinkin kautta, 5'.
Voi palvella sinua: kodonia- RNM: n 3 '-pää kärsii "klivaje" endonukleaasilla, joka tuottaa vapaan hydroksyyliryhmän 3', joka liittyy adeniinijätteen "ristraan" tai "hännän" (100 - 250), jotka lisätään samaan aikaan entsyymin aika Poly (A) polymeraasi.
"Hood 5" "ja" häntä Poly A ”he suorittavat toiminnot RNM -molekyylien suojaamisessa hajoamiselta ja lisäksi ne toimivat kypsien transkriptien kuljettamisessa sytosoliin ja vastaavasti translaation aloittamisessa ja lopettamisessa.
COrte ja Empalme
Transkription jälkeen "ensisijaiset" RNM: t, joissa on kaksi modifioitua äärimmäisyyttään, jotka ovat edelleen läsnä ytimessä, kulkevat "leikkaus- ja silmukointi" -prosessin läpi, jolla introniikkisekvenssit yleensä eliminoidaan ja tuloksena olevat eksonit on liitetty (alueellisen prosessoinnin jälkeinen prosessi), Mitä kypsät transkriptit, jotka luopuvat ytimestä ja saavuttavat sytosolin.
Leikkaus ja silmukointi suoritetaan riboprotic -kompleksilla, nimeltään Espicleosoma (Anglismi Silmukka), muodostettu viisi pientä ribonukleoproteiinia ja RNA -molekyyliä, jotka kykenevät "tunnistamaan" alueet, jotka on poistettava primaarisesta transkriptista.
Monissa eukaryooteissa on ilmiö, joka tunnetaan nimellä "leikkaus ja vaihtoehtoinen nivel", mikä tarkoittaa, että erityyppiset rekisteröintien jälkeiset modifikaatiot voivat aiheuttaa erilaisia tai isoentsyymiproteiineja, jotka eroavat toisistaan tietyillä sekvenssien osa -alueilla.
- Ribosomit
Kun kypsät transkriptit poistuvat ytimestä ja kuljetetaan translaatioon sytosoliksi, ne prosessoivat RNA -molekyyleihin liittyvästä proteiinikompleksista tunnetulla translaatiokompleksella, joka koostuu ribosomiksi, joka koostuu proteiinikompleksista.
Ribosomit koostuvat kahdesta alayksiköstä, yhdestä "suuresta" ja toisesta "pienestä", jotka ovat vapaasti dissosioituneita sytosolissa ja yhdistyvät tai yhdistävät mRNA -molekyylin.
Ribosomien ja mRNA: n välinen liitto riippuu erikoistuneista RNA -molekyyleistä, jotka liittyvät ribosomaalisiin proteiineihin (ribosomaalinen RNA tai RNA ja siirto tai ARNT RNA), joista kukin harjoittaa spesifisiä toimintoja.
ARNT ovat molekyyli "adapterit", koska yhden päätään he voivat "lukea" jokaisen kodonin tai kolminkertaisen kypsän RNA: n (emäksen komplementaarisesti) ja toisen kautta ne voivat liittyä kodonin koodaavaan aminohappohapuun "lue" "lue" "lue" "lukema".
Toisaalta RNR -molekyylit ovat vastuussa kunkin aminohapon sitoutumisprosessin kiihdyttämisestä syntyvässä peptidiketjussa.
Eukaryoottinen kypsä RNM voidaan "lukea" monilla ribosomeilla, niin monta kertaa kuin solu osoittaa sen. Toisin sanoen sama RNM voi johtaa moniin kopioihin samasta proteiinista.
Kodonin lähtö- ja lukukehys
Kun ribosomaaliset alayksiköt lähestyvät kypsää RNM: ää, riboproottinen kompleksi "skannaa" mainitun molekyylin sekvenssin, kunnes aloituskodonia löytyy, joka on aina AUG ja tarkoittaa metioniinitähteen käyttöönottoa.
Se voi palvella sinua: Monoploidy: Kuinka se tapahtuu, organismit, taajuus ja hyödyllisyysAUG -kodonin määrittelee kunkin geenin lukukehyksen ja määrittelee lisäksi kaikkien luonnon transloidujen proteiinien ensimmäisen aminohapon (tämä aminohappo eliminoidaan usein translaation jälkeen).
Päätekodonit
Kolme muuta kodonia on tunnistettu sellaisiksi, jotka indusoivat käännöksen lopettamisen: UAA, UAG ja UGA.
Ne mutaatiot, jotka tarkoittavat aminohappoa koodaavan tripletin typpimuutoksia ja jotka johtavat päättymiskodoneihin, tunnetaan merkityksettöminä mutaatioina, koska ne aiheuttavat synteesiprosessin ennenaikaisen pidätyksen, joka muodostaa lyhyemmät lyhyemmät proteiinit.
Kääntämättömät alueet
Lähellä kypsien RNM -molekyylien 5 'päätä on alueita, joita ei käännetä (UTR, englannista Irrottaa alue), nimeltään myös ”johtaja” -sekvenssejä, jotka sijaitsevat ensimmäisen nukleotidin ja translaation alkamisen välillä (AUG).
Näillä UTR -alueilla, joita ei käännetä.
- Käännöksen aloitus
Käännös, samoin kuin transkriptio koostuu kolmesta vaiheesta: yksi aloituksesta, toinen pidentymisestä ja lopulta yksi lopettaminen.
Aloitus
Se koostuu RNM: n translaatiokompleksin kokoonpanosta, joka ansaitsee kolmen proteiinin yhdistyksen, joka tunnetaan nimellä aloituskertoimet (jos, englanninkielisiä Aloituskerroin) If1, if2 ja if3 ribosomin pieneen alayksikköön.
Aloituskertoimien ja pienen ribosomaalisen alayksikön muodostama "pre -jaeaatiota" -kompleksi liitetään puolestaan, ja Arnt, joka "kuormittaa" metioniinitähdettä ja tämä molekyylijoukko sitoutuu RNAM: iin, lähellä lähtökodonia AUG, AUG.
Nämä tapahtumat johtavat RNM -liittoon suuren ribosomaalisen alayksikön kanssa, mikä johtaa aloitustekijöiden vapauttamiseen. Ribosomin suurella alayksiköllä on 3 Union -kohtaa ARNT -molekyyleille: kohta A (aminohappo), kohta P (polypeptidi) ja kohta E (lähtö).
Sivusto A liittyy aminoacil-Arnt-antikodiin, joka on täydentävä käännetyn mRNA: n kanssa; P -paikka on aminohappo siirretty ARNT: stä syntyvään peptidiin ja S -kohta on siellä, missä se on Arnt “tyhjä” ennen kuin se vapautuu sytosoliin aminohapon toimittamisen jälkeen.
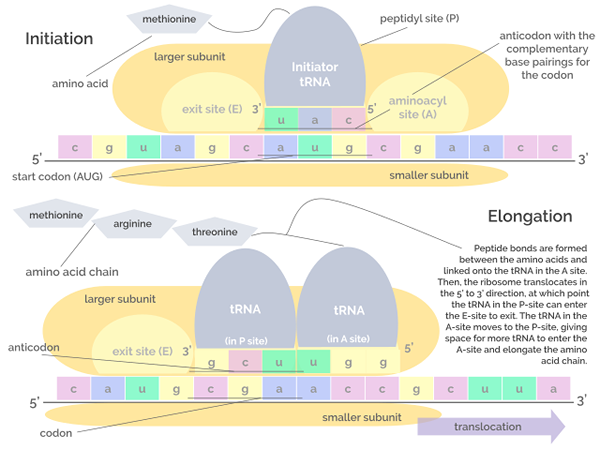 Käännöksen aloitus- ja pidentymisvaiheiden graafinen esitys (lähde: Jordan Nguyen/CC BY-S (https: // creativecommons.Org/lisenssit/by-SA/4.0) Wikimedia Commonsin kautta)
Käännöksen aloitus- ja pidentymisvaiheiden graafinen esitys (lähde: Jordan Nguyen/CC BY-S (https: // creativecommons.Org/lisenssit/by-SA/4.0) Wikimedia Commonsin kautta) Pidennys
Tämä vaihe koostuu ribosomin "liikkeestä" mRNA -molekyyliä pitkin ja kunkin kodonin translaatio, joka "lukee", mikä merkitsee polypeptidiketjun kasvua tai pidentymistä syntymässä.
Tämä prosessi vaatii GTP: n pidentymiskerroksena tunnetun tekijän GTP: n muodossa, joka ajaa pidentymistekijöiden siirtymistä RNM -molekyyliä pitkin sen aikana.
Voi palvella sinua: Okazaki -fragmentitRibosomaalisten RNA: ien transferaasipeptidiliaktiivisuus mahdollistaa peptidisidosten muodostumisen ketjuun lisättyjen aminohappojen välillä.
Irtisanominen
Käännös päättyy, kun ribosomi kohtaa minkä tahansa lopetuskodonin, koska ARN: t eivät tunnista näitä kodoneja (ne eivät koodaa aminohappoja). Yhdistetään myös proteiinit, jotka tunnetaan myös vapautumiskertoimina, mikä helpottaa ribosoma MR: n irrottamista.
Procary-käännös (passes-prosessit)
Prokaryooteissa, kuten eukaryoottisoluissa, proteiinisynteesin varausrbosomit löytyvät sytosolista (mikä pätee myös transkriptionaalisiin koneisiin), tosiasia, joka mahdollistaa proteiinin sytosolisen pitoisuuden nopean lisääntymisen geenien ilmentymisen yhteydessä, jotka geenien ekspressio koodaa se kasvaa.
Vaikka transkription aikana tuotettu ensisijainen RNM ei ole erittäin yleinen prosessi näissä organismeissa, rekisteröinnin jälkeinen kypsyminen voi kärsiä "leikkaamisen ja silmukoinnin" kautta. Yleisin on kuitenkin tarkkailla ensisijaiseen transkriptoituun ribosomeihin, jotka kääntävät sen samanaikaisesti, kun se transkriptoidaan vastaavasta DNA -sekvenssistä.
Edellä esitetyn kannalta monien prokaryootien käännös alkaa 5: llä ', koska mRNA: n 3' pää pysyy kytkettynä muotti -DNA: han (ja tapahtuu samanaikaisesti transkription kanssa).
Kääntämättömät alueet
Prokaryoottiset solut tuottavat myös RNM: n kääntämättömien alueiden kanssa, jotka tunnetaan nimellä "Shine-Dalgarno Box" ja joiden konsensussekvenssi on Aggagg. Kuten on selvää, bakteerien UTR -alueet ovat huomattavasti lyhyempiä kuin eukaryoottisoluissa, vaikka ne käyttävät samanlaisia toimintoja translaation aikana.
Käsitellä asiaa
Bakteereissa ja muissa prokaryoottisissa organismeissa translaatioprosessi on melko samanlainen kuin eukaryoottiset solut. Se koostuu myös kolmesta vaiheesta: aloittaminen, pidentyminen ja lopettaminen, jotka riippuvat erityisistä prokaryoottisista tekijöistä, jotka eroavat Eukaryootien käyttämistä.
Esimerkiksi pidennys riippuu eukaryoottisen tekijän G sijasta tunnetuista pidentymistekijöistä, jotka tunnetaan nimellä ef-tu ja ef-ts.
Viitteet
- Alberts, b., Johnson, a., Lewis, J., Raff, m., Roberts, k., & Walter, P. (2007). Solumolekyylin biologia. Garlantitiede. New York, 1392.
- Savi, s. & Brown, W. (2008) Translaatio: DNA mRNA: lle proteiiniksi. Luontokoulutus 1 (1): 101.
- Griffiths, a. J -., Wessler, S. R -., Lewontin, R. C., Gelbart, W. M., Suzuki, D. T., & Miller, J. H. (2005). Johdatus geenianalyysiin. Macmillan.
- Loodish, h., Berk, a., Kaiser, c. -Lla., Krieger, m., Scott, M. P., Bretscher, a.,… & Matsudaira, P. (2008). Molekyylisolubiologia. Macmillan.
- Nelson, D. Lens., Lehninger, a. Lens., & Cox, M. M. (2008). Lehninger -biokemian periaatteet. Macmillan.
- Rosenberg, L. JA., & Rosenberg, D. D -d. (2012). Ihmisen geenit ja genomit: Tiede. Terveys, yhteiskunta, 317-338.
- « Sorbus Aria -ominaisuudet, elinympäristö, ominaisuudet, kulttuuri
- Boletus aereus -ominaisuudet, elinympäristö, tunnistaminen, reseptit »