

Urasiilirakenne, toiminnot, ominaisuudet, synteesi
- 1016
- 69
- Sheldon Kuhn
Hän Urasiili Se on pyrimidiinin typpipohja, jota löytyy ribonukleiinihaposta (RNA). Tämä on yksi ominaisuuksista, jotka erottavat RNA: n deoksiribonukleiinihaposta (DNA), koska jälkimmäisellä on Timina urasiilin sijasta. Molemmat aineet, urasiili ja Timina, vain eroavat siitä, että toisella on metyyliryhmä.
Evoluutio -kannalta on ehdotettu, että RNA oli ensimmäinen molekyyli, joka säilytti geneettisen tiedon ja toimi katalyyttinä soluissa, ennen DNA: ta ja entsyymejä. Tämän vuoksi ajatellaan, että urasiililla oli avainrooli elämän kehityksessä.
Lähde: Kemikungen [julkinen alue] Elävissä olennoissa urasiilia ei löydy vapaasti, vaan yleisesti monofosfaattinukleotideja (UMP), difosfaattia (UDP) ja trfosfaattia (UTP). Näillä urasiilinukleotideilla on erilaiset toiminnot, kuten RNA -biosynteesi ja glykogeeni, sokerien isomeerinen käännös ja syntaasin syntaasin säätely.
[TOC]
Rakenne ja ominaisuudet
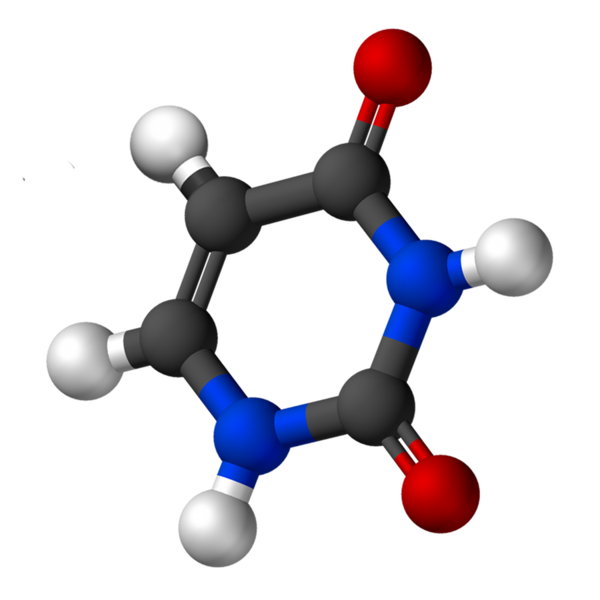
Urasiililla, jota kutsutaan 2,4-dioksipiridiiniksi, on empiirinen kaava C4H4N2JOMPIKUMPI2, jonka molekyylipaino on 112,09 g/mol ja puhdistetaan valkoisena jauheena.
Uridiinin rakenne on heterosyklinen rengas, jossa on neljä hiiliatomia ja kaksi typpeä, kaksoisvaihtoehtoisilla sidoksilla. Se on tasomainen.
Sillä on 50 mg/ml liukoisuus, 25 ºC, 1 m natriumhydroksidissa ja PKA 7: n välillä.9 ja 8.2. Aallonpituus, jossa sen suurin absorbanssi tapahtuu (ʎMax) Se on välillä 258 - 260 nm.
Biosynteesi
Pyrimidiininukleotidien (urasiili ja sytokiini) biosynteesille on yleinen polku. Ensimmäinen askel on co: n karbamoilifosfaatin biosynteesi2 ja NH4+, jota katalysoi karbamoilisyntetaasifosfaatti.
Pyrimidiini on rakennettu hiilihiilihallinnasta ja aspartaatista. Molemmat aineet reagoivat ja muodostavat N-karbamoilaspartato, reaktio, jota katalysoi transcabamoilasa-aspartaatti (ATCASA). Pyrimidiinirenkaan sulkeminen voidaan dehydratoitua dihydrootaasilla, ja se tuottaa L-dihydrotatoaatin.
L-dihydrotoaatti hapetetaan ja muuttuu orotateiksi; Elektronia vastaanottaja on nad+. Se on reaktio, jota katalysoi dihydroorotaattidehydrogenaasi. Seuraava vaihe koostuu fosforibosyyliryhmän siirrosta fosforibosyylipyrofosfaatista (PRPP). Gradidilato -muoto (WPO) ja epäorgaaninen piroffosfaatti (PPI), katalysoimalla fosforibosyyliorotaatin transferaasilla.
Viimeinen vaihe koostuu ryhmän pyrimidiinirenkaan (WPO) dekarboksyloinnista. Uridilaato-muoto (uridiini-5'-monofosfaatti, UMP), jota katalysoidaan dekarboksylaasilla.
Sitten kinaasin osallistumisen kautta fosfaattiryhmä siirretään ATP: stä UMP: hen, muodostaen UDP: n (uridiini-5'-difosfaatti). Jälkimmäinen toistetaan, muodostaen UTP: n (uridin-5'-triffosfaatti).
Voi palvella sinua: biologian haarat ja mitä he opiskelevatBiosynteesin säätely
Bakteereissa pyrimidiinibiosynteesin säätely tapahtuu negatiivisen palautteen avulla, TransCabamoilasa -aspartaattitasolla (ATCASA).
Tätä entsyymiä estää CTP (citidin-5'-trofosfaatti), joka on pyrimidiinien biosynteesireiän lopullinen tuote. Atcas.
Eläimissä pyrimidiinin biosynteesin säätely tapahtuu negatiivisen palautteen avulla kahden entsyymin tasolla: 1) karbamoilisyntaasi II -fosfaatti, jota UTP estää ja aktivoi ATP ja PRPP; ja 2) OMP -diskarboksylaasi, jota estyy itse katalysoidun reaktion tuote, UMP. OMP: n biosynteesin nopeus vaihtelee PRPP: n saatavuuden mukaan.
Toiminta RNA: n biosynteesissä
Urasiili on läsnä kaikentyyppisissä RNA: ssa, kuten Messenger -RNA (MNA), siirto -RNA (ARNT) ja ribosomaalinen RNA (RNA). Näiden molekyylien biosynteesi tapahtuu prosessin kautta, jota kutsutaan transkriptioksi.
Transkription aikana DNA: n sisältämät tiedot kopioidaan RNA: ssa pon -polymeraasi -RNA: n kautta. Käänteinen prosessi, jossa RNA: n tiedot kopioidaan DNA: ssa, tapahtuu joissakin viruksissa ja kasveissa käänteistranskriptaasin läpi.
RNA -biosynteesi tarvitsee tryposfaattinukleosideja (NTP), nimittäin: tryfosfaatti uridiini (UTP), tryposfaattisyidiini (CTP), adeniinin tryposfaatti -adeniini (ATP) ja tryfosfaatti -guaniini (GTP). Reaktio on:
(RNA)n jätteet + NTP -> (RNA)N+1 jäte + PPI
Epäorgaaninen pyrofosfaatti (PPI) -hydrolyysi tarjoaa energiaa RNA: n biosynteesille.
Toiminto sokerin biosynteesissä
Sokeresterit ovat hyvin yleisiä elävissä organismeissa. Jotkut näistä estereistä ovat nukleosidiesterien, kuten UDP-sokerin, difosfas, jotka ovat hyvin runsaasti soluissa. UDP-sokeri sisältää disakkaridien, oligosakkaridien ja polysakkaridien biosynteesiä.
Kasveissa sakkaroosin biosynteesi tapahtuu kahdella tavalla: pää- ja toissijainen.
Pääreitti koostuu D-glukoosin siirtämisestä UDP-D-glukosasta D-frucease-sakkaroosin ja UDP: n muodostamiseksi. Toissijainen tie sisältää kaksi vaihetta: se alkaa UDP-D-glukoosilla ja fruktoosi-6-fosfaatilla ja huipentuu sakkaroosin ja fosfaatin muodostumisen kanssa.
Myrskyrauhasissa laktoosibiosynteesi tapahtuu UDP-D-galaktoosista ja glukoosista.
Voi palvella sinua: biokemian haaratKasveissa selluloosabiosynteesi suoritetaan beeta-d-glykosyylijätteiden jatkuvalla kondensaatiolla UDP-glukoosista kasvavan polyglukoosiketjun vähentämättömään päähän. Samoin amyloosi- ja amylopektiini-biosynteesi vaativat UDP-glukoosin glukoosin luovuttajien substraattina kasvavalle ketjulle.
Eläimissä käytetään sekä UDP-glukoosia että ADP-glukoosia glykogeenin biosynteesiin. Samoin kondroitiinisulfaattibiosynteesi vaatii UDP-ksyloosin, UDP-galaktoosia ja UDP-glucuronatoa.
Toiminto sokerien isomeerisessä käännöksessä
Galaktoosin muuntaminen glykolyysin välittäjäksi tapahtuu Leloir -tien kautta. Yhden tämän reitin vaiheista katalysoi UDP-galaktoosi-4-epimeraasientsyymi, joka helpottaa UDP-galaktoosin välistä kääntyä UDP-glukoosiin.
Glykoproteiinien biosynteesissä
Glykoproteiinien biosynteesin aikana proteiinit kulkevat Golgi -laitteen cis-, väliaine- ja trans -säkkien läpi.
Jokaisessa näistä laukkuista on joukko entsyymejä, jotka käsittelevät glykoproteiineja. Sokerimonomeerit, kuten glukoosi ja galaktoosi.
Nukleotidi-heksoosi kuljetetaan Golgi-säiliöihin antiporten avulla. UDP-galaktoosi (UDP-GAL) ja UDP-N-asetyyligalaktosamina (UDP-GALNAC) tulevat säiliöihin sytosolista vaihtamisella UMP: llä.
Golgi -säiliössä fosfataasi hydrolysoi UDP- ja UMP- ja PI -fosfaattiryhmää. UDP tulee reaktioista, joita katalysoivat galaktosyylitransferaasi ja N-asetyyligalaktosamiltransferaasi. Fosfataasilla muodostettu UMP palvelee nukleotidien-heksoosin vaihtamista.
Toiminto syntaasin säätelyssä
Mekanismi glutamiinisyntaasin säätelemiseksi on kovalenttinen modifikaatio, joka koostuu adenilaatiosta, joka on inaktiivinen ja kylki, jotka aktiivisesti se. Tämä kovalenttinen modifikaatio on palautuva ja katalysoi adenyylitransferaasi.
Adenyylitransferaasin aktiivisuutta moduloi PII -proteiinin liitolla, jota säätelee kovalenttinen modifikaatio, uridinilaatio.
Sekä uridilililaation että siirtymän suorittaa uridililtransferaasi. Tässä entsyymissä uridilaation aktiivisuus johtuu glutamiinista ja fosfaatista, ja sen aktivoidaan alfa-zotoglutaraatin ja ATP: n liitolla PII: hen.
Toiminto RNA -painos
Joitakin RNM: ää muokataan ennen käännöstä. Joissakin eukaryoottisissa organismeissa, kuten Trypanosoma Brucei, Sytokromioksidaasin alayksikön II geenin transkriptin RNA on painos. Näin tapahtuu asettamalla urasiilijätteet, terminaali uridiltransferaasi katalysoima reaktio.
Voi palvella sinua: Curl: Koostumus, osat, merkitysMuokatun tuotteen täydentävä opas RNA toimii maltillisena muokkausprosessissa. Alkuperäisen transkriptin ja ohjaus-RNA: n välillä muodostetut emäsparit viittaavat emäspariin g = u, jotka eivät ole Watson-Record ja ovat yleisiä RNA: ssa.
UDP-glukoosibiosynteesi
Fysiologisissa olosuhteissa glykogeenin biosynteesi glukoosi-1-fosfaatista on termodynaamisesti mahdotonta (positiivinen AG). Tämän vuoksi ennen biosynteesiä tapahtuu glukoosi-1-fosfaatin (G1P) aktivointi. Tämä G1P ja UTP yhdistää reaktion glukoosidifosfaatin uridiinin (UDP-glukoosin tai UDPG) muodostamiseksi.
Reaktiota katalysoi UDP-glukoosin pyrofosforyillaasi, ja se on seuraava:
G1P + UTP -> udp -glucosa + 2pi.
Gibbs-vapaa energian vaihtelu tässä vaiheessa on suuri ja negatiivinen (-33,5 kJ/mol). Happireaktion aikana G1P hyökkää UDP-glukoosin ja epäorgaanisen pyrofosfaatin (PPI) Alforo Alpha -fosforiatomiin. Sitten epäorgaaniset pyrofosfaatit hydrolysoivat PPI: tä, jonka hydrolyysienergia on se, joka ajaa yleistä reaktiota.
UDP-glukoosi on "korkean energian" aine. Se mahdollistaa glykosidisidosten muodostamisen glukoositähteen ja kasvavan polysakkaridiketjun välillä. Sama energiaperiaate voidaan soveltaa reaktioihin, joissa UDP-sokeri osallistuu, kuten disakkaridien, oligosakkaridien ja glykoproteiinien biosynteesi.
Urasiili -DNA Glycosilasa
On DNA -vaurioita, jotka tapahtuvat spontaanisti. Yksi näistä vaurioista on sprontaani. Tässä tapauksessa korjaus tapahtuu modifioidun DNA -emäksen takia entsyymin avulla, jota kutsutaan Urasiili -DNA -glycosilasaksi.
Entsyymi Urasiili-DNA-glykosilasa eliminoi vaurioituneen sytokiinin (urasiili), joka tuottaa deoksihiomajäännöstä, josta puuttuu typpipohja, nimeltään AP-alue (Apurinic-Apirimidiininen kohta).
Sitten endonukleaasi-AP-entsyymi tekee leikkauksen AP-kohdan fosfodiesterin luurankoon eliminoimalla sokerifosfaattijäämä. DNA -polymeraasi I palauttaa vaurioituneen juosteen.
Viitteet
- Bohinski, r. 1991. Biokemia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokemia. Toimitusverkko, Barcelona.
- Loodish, h., Berk, a., Zipurski, S.Lens., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Solu- ja molekyylibiologia. Toimitus Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Meksiko, Sāo Paulo.
- Nelson, D. Lens., Cox, m. M. 2008. Biokemian lehninger-periaatteet. W -.H. Freeman, New York.
- Voet, D. ja Voet, J. 2004. Biokemia. John Wiley ja pojat, USA.
- « Marginaalikustannuskaava, laskenta ja harjoitukset
- Pyrimidiinien ominaisuudet, rakenne, toiminnot »