

Ketogeneesityypit, synteesi ja hajoaminen
- 3177
- 113
- Ronald Reilly
Se ketogeneesi Se on prosessi, jolla saadaan asetoasetaatti, β-hydroksibutiraatti ja asetoni, joita yhdessä kutsutaan ketonirunkoiksi. Tämä monimutkainen ja hienoksi säännelty mekanismi suoritetaan mitokondrioissa rasvahappojen katabolismista.
Ketonirunkojen hankkiminen tapahtuu, kun vartalo on tyhjentäviä paastojaksoja. Vaikka nämä metaboliitit syntetisoidaan enimmäkseen maksasoluissa, niitä havaitaan tärkeänä energianlähteenä monimuotoisissa kudoksissa, kuten luurankojen lihaksissa ja sydämessä ja aivokudoksissa.
Lähde: SAV VAS [CC0] Β-hydroksibutiraatti ja asetoasetaatti ovat metaboliitteja, joita käytetään substraattina sydänlihaksessa ja munuaiskuoressa. Aivoissa ketonirunkoista tulee tärkeitä energialähteitä, kun vartalo on käyttänyt glukoosireseriä.
[TOC]
Yleiset luonteenpiirteet
Ketogeneesiä pidetään erittäin tärkeänä fysiologisena tai aineenvaihduntareitin. Yleensä tämä mekanismi suoritetaan maksassa, vaikkakin on osoitettu, että se voidaan suorittaa muissa kudoksissa, jotka kykenevät metaboloimaan rasvahappoja.
Ketonirunkojen muodostuminen on asetyyli-CoA: n tärkein metabolinen johdannainen. Tämä metaboliitti saadaan metabolisesta reitistä, joka tunnetaan nimellä β-hapettuminen, joka on rasvahappojen hajoaminen.
Glukoosin saatavuus kudoksissa, joissa β-hapettuminen tapahtuu, määrittää asetyyli-CoA: n metabolisen kohteen. Erityisesti hapettuneet rasvahapot kohdistuvat melkein kokonaan ketonirunkojen synteesiin.
Ketonirunkojen tyypit ja ominaisuudet
Tärkein ketoninen runko on asetoasetaatti tai asetoetikkahappo, joka syntetisoidaan enimmäkseen maksasoluissa. Asetoasetaatista muut ketonirungot muodostavat molekyylit ovat johdettu.
Asetoetikkahapon pelkistys aiheuttaa D-β-hydroksibutiraatin, toisen ketonisen kehon. Asetoni on vaikea yhdiste hajoamiseen, ja sitä tuotetaan asetoasetaatin spontaanilla dekarboksylaatioreaktiolla (joten se ei vaadi minkään entsyymin puuttumista), kun sitä on suurissa pitoisuuksissa veressä.
Ketonirunkojen nimellisarvo on järjestetty yleissopimuksella, koska tiukasti β-hydroksibutiraatilla ei ole ketonitoimintoa. Nämä kolme molekyyliä liukenevat veteen, mikä helpottaa niiden verenkuljetusta. Sen päätehtävä on tarjota energiaa tietyille kudoksille luuston ja sydämen lihaksena.
Ketonirunkojen muodostumiseen osallistuvat entsyymit ovat pääasiassa maksassa ja munuaisissa, mikä selittää, että nämä kaksi sijaintia ovat näiden metaboliittien tärkeimmät tuottajat. Sen synteesi esiintyy vain ja yksinomaan solujen mitokondriaalimatriisissa.
Se voi palvella sinua: itiö: kasveissa, sienissä ja bakteereissaKun nämä molekyylit on syntetisoitu, ne menevät verenkiertoon käsittelemällä niitä vaativia kudoksia, joissa ne hajoavat, kunnes asetyyli-CoAA.
Ketonirunkojen synteesi
Ketogeneesin olosuhteet
Asetyyli-CoA: n metabolinen kohde β-hapettumisesta riippuu organismin metabolisista vaatimuksista. Tämä hapetetaan CO: lle2 ja h2Tai sitruunahapposyklin tai rasvahappojen synteesin avulla, jos lipidien ja hiilihydraattien metabolia on stabiili kehossa.
Kun vartalo tarvitsee hiilihydraatteja, oksalasetaattia käytetään glukoosinvalmistukseen (glukoneogeneesi) sitruunahapposyklin käynnistämisen sijasta. Tämä tapahtuu, kuten mainittiin, kun keholla on jonkin verran kyvyttömyyttä saada glukoosia, esimerkiksi pitkäaikaisessa paastossa tai diabeteksen läsnäolossa.
Tästä johtuen rasvahappojen hapettumisesta johtuva asetyyli-CoA käytetään ketonirunkojen tuotantoon.
Mekanismi
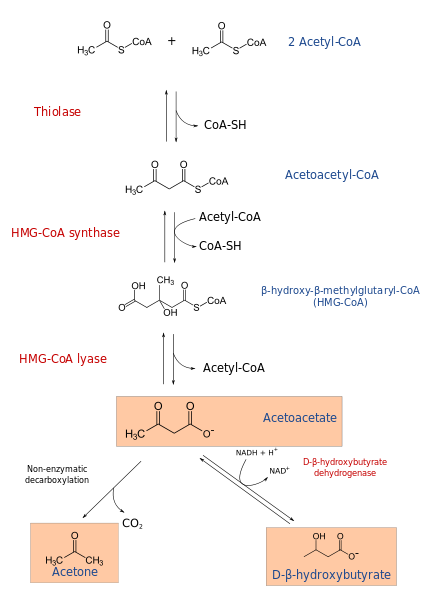
Ketogeneesiprosessi alkaa β-hapettumisen tuotteista: asetasetyyli-CoA tai asetyyli-CoA. Kun substraatti on asetyyli-CoA, ensimmäinen vaihe koostuu kahden molekyylin kondensaatiosta, asetyyli-CoA-transferaasireaktiosta, asetatyyli-CoA: n tuottamiseksi.
Asetacetyyli-CoA tiivistetään kolmannella asetyyli-CoA: lla HMG-CoA-syntaasin vaikutuksella HMG-CoA: n (β-hydroksi-β-metyyliglutariil-CoA) tuottamiseksi (β-hydroksi-β-metyyliglutariil-CoA). HMG-CoA on hajotettu asetoasetaattiin ja asetyyli-CoA: ksi HMG-CoA Liasa -toimella. Tällä tavoin saadaan ensimmäinen ketonikeho.
Asetoasetaatti pelkistetään p-hydroksibutiraatiksi beeta-hydroksibutiraattidehydrogenaasin interventiolla. Tämä reaktio riippuu NADH: sta.
Ketoninen asetoasetaattirunko on β-pethoacid, joka kokee ei-entsymaattisen dekarboksylaation. Tämä prosessi on yksinkertainen ja tuottaa asetonin ja yhteistyökumppanin2.
Tämä reaktiosarja johtaa siten ketonirunkoihin. Ne, jotka liukenevat veteen.
Β-hapettuminen ja ketogeneesi liittyvät
Rasvahappojen aineenvaihdunta tuottaa substraatteja ketogeneesille, joten nämä kaksi tapaa ovat toiminnallisesti liittyvät.
Asetoaketyyli-CoA on rasvahappojen metabolian estäjä, koska se pysäyttää asyyli-CoA-dehydrogenaasin aktiivisuuden, joka on β-hapettumisen ensimmäinen entsyymi. Lisäksi se harjoittaa myös estämistä asetyyli-CoA -siirtoon ja HMG-CoA-syntaasiin.
HMG-CoA-syntaasin entsyymi, jollei CPT-I (entsyymi, joka liittyy asyylikarnitiinin tuotantoon β-hapettumisessa), edustaa tärkeätä säätelevää roolia rasvahappojen muodostumisessa.
Se voi palvella sinua: Zacatecasin kasvisto ja eläimistö: edustavampi lajiΒ-hapettumisen säätely ja sen vaikutus ketogeneesiin
Organismin ruokinta säätelee monimutkaista sarjaa hormonaalisia signaaleja. Ruokavaliossa kulutetut hiilihydraatit, aminohapot ja lipidit kerrostuvat triasyyliglyserolien muodossa rasvakudoksessa. Insuliini, anabolinen hormoni, puuttuu lipidien synteesiin ja triakyyliglyserolien muodostumiseen.
Mitokondrioiden tasolla β-hapettumista säätelee joidenkin substraattien pääsy ja osallistuminen mitokondrioihin. Entsyymi CPT I syntetisoi karnitiiniasyylia ACIL -sytosolisesta ACIL: stä.
Kun kehoa syötetään, asetyyli-CoA-karboksylaasi ja sitraatti lisää CPT I: n tasoja, samalla kun se vähentää sen fosforylaatiota (syklinen AMP-riippuvainen reaktio).
Tämä aiheuttaa Malonil COA: n kertymisen, joka stimuloi rasvahappojen synteesiä ja estää sen hapettumisen estäen turhaa syklin syntymistä.
Paaston tapauksessa karboksylaasin aktiivisuus on erittäin alhainen, koska entsyymin CPT: n tasot ovat vähentyneet ja on myös fosforyloitunut, aktivoimalla ja edistämällä lipidien hapettumista, mikä myöhemmin sallii ketonirunkojen muodostumisen asetyylien läpi -Coa.
Heikkeneminen
Ketonirungot levisivät solujen ulkopuolelle, joissa ne syntetisoitiin ja verenkierto kuljetetaan perifeerisiin kudoksiin. Näissä kudoksissa ne voidaan hapettaa trikarboksyylihappojen syklin kautta.
Perifeerisissä kudoksissa β-hydroksibutiraatti hapetetaan asetoasetaattiin. Myöhemmin nykyinen asetoasetaatti aktivoidaan 3-ZOA-transferaasientsyymin vaikutuksella.
Succinil-CoA toimii COA-luovuttajana tulossa sukkinaatiksi. Asetoasetaatin aktivointi tapahtuu sukkinyyli-CoA: n estämiseksi.
Tuloksena oleva aceoasetyyli-CoA kärsii tioliittisesta repeämästä, joka tuottaa kaksi asetyyli-CoA-molekyyliä, jotka sisällytetään trikarboksyylihappojen sykliin, tunnetaan paremmin nimellä Krebs-sykli.
Maksasoluista puuttuu 3-kotoasil-CoA-siirto, joka estää tämän metaboliitin aktivoinnin näissä soluissa. Tällä tavalla on taata, että ketonirungot eivät hapettele soluissa, joissa ne tuotetaan, mutta että ne voidaan siirtää kudoksiin, joissa niiden aktiivisuus vaaditaan.
Ketonirunkojen lääketieteellinen merkitys
Ihmiskehossa veressä olevat korkeat ketonirunkojen pitoisuudet voivat aiheuttaa erityisiä olosuhteita, joita kutsutaan asidoosiksi ja ketonemiaksi.
Voi palvella sinua: Sphingomyeline: Mikä on, rakenne, toiminnot, synteesiNäiden metaboliittien valmistus vastaa rasvahappojen ja hiilihydraattien katabolismia. Yksi yleisimmistä patologisen ketogeneesin tilan syistä on etikkarikarbonoitujen fragmenttien korkea pitoisuus, jotka eivät hajoa trikarboksyylihappojen hapettumisreittiä.
Seurauksena on,. Tämä tarkoittaa näiden metaboliittien välittäjäaineenvaihdunnan häiriöitä.
Tietyt aivolisäkkeen neuroGandular -tekijöiden puutteet, jotka säätelevät ketonirunkojen hajoamista ja synteesiä yhdessä hiilivetyjen aineenvaihdunnan häiriöiden kanssa, ovat syynä hyperketonemian tilaan.
Diabetes mellitus ja ketonikehojen kertyminen
Diabetes mellitus (tyyppi 1) on endokriininen sairaus, joka aiheuttaa ketonirunkojen tuotannon lisääntymisen. Insuliini riittämätön tuotanto poistaa glukoosin lihaksiin, maksaan ja rasvakudokseen, joten veressä kertyy.
Solut glukoosin puuttuessa alkavat glukoneogeneesin ja rasvan ja proteiinien hajoamisen prosessin niiden aineenvaihdunnan palauttamiseksi. Seurauksena on, että oksalasetaattipitoisuudet vähenevät ja lisäävät lipidien hapettumista.
Asetyyli-CoA: n kertyminen tapahtuu, joka oksalastaatin puuttuessa ei voi seurata sitruunahappoa, aiheuttaen sitten ketonirunkojen korkeat tuotannot, jotka ovat ominaisia tälle sairaudelle.
Asetonin kertyminen havaitaan sen läsnäolon virtsassa ja hengityksessä, jotka esittävät tämän tilan, ja on itse asiassa yksi oireista, jotka osoittavat tämän taudin ilmenemisen.
Viitteet
- Blázquez ortiz, c. (2004). Ketogeneesi astrosyyteissä: karakterisointi, säätely ja mahdollinen sytoprotektiivinen paperi (Väitöskirjan tohtori, Computense University of Madridin yliopisto, Publications Service).
- Devlin, t. M. (1992). Biokemian oppikirja: kliinisillä korrelaatioilla.
- Garrett, r. H., & Grisham, c. M. (2008). Biokemia. Thomson Brooks/Cole.
- McGry, J. D -d., Mannaerts, g. P., & Foster, D. W -. (1977). Malonyyli-CoA: n mahdollinen rooli maksan rasvahappojen hapettumisen ja ketogeneesin säätelyssä. Journal of Clinical Investigation, 60(1), 265-270.
- Melo, v., Ruiz, v. M., & Cuamatzi tai. (2007). Aineenvaihduntaprosessien biokemia. Palautus.
- Nelson, D. Lens., Lehninger, a. Lens., & Cox, M. M. (2008). Lehninger -biokemian periaatteet. Macmillan.
- Pertierra, a. G., Gutiérrez, c. V., & Muut, c. M. (2000). Aineenvaihdunnan biokemian perusteet. Toimitus Tébar.
- Voet, D., & Voet, J. G. (2006). Biokemia. Ed. Pan -American Medical.
- « Mannermainen aamiainen mikä on ja mitä ruokia se sisältää?
- Kalsiumperoksidi (CAO2) ominaisuudet, riskit ja käytöt »