

Kolme ominaisuutta ja toiminto organismissa
- 968
- 188
- Arthur Terry II
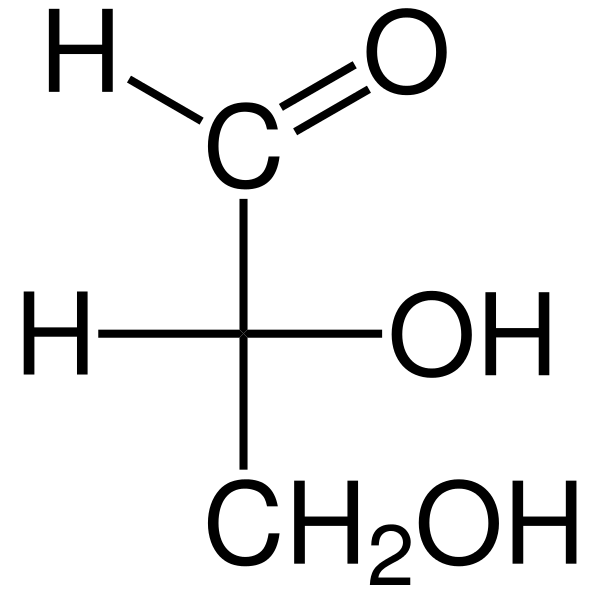
Se Triosas Ne ovat kolmen hiilen monosakkarideja, joiden empiirinen kemiallinen kaava on c3H6JOMPIKUMPI6. On kaksi triosaa: glykaraldehyd. Triosat ovat tärkeitä aineenvaihdunnassa, koska ne yhdistävät kolme metabolista reittiä: glykolyysi, glukoneogeneesi ja pentoosifosfaattipolku.
Fotosynteesin aikana Calvin-sykli on triosan lähde, jotka palvelevat fruktoosi-6-fosfaattien biosynteesiä. Tämä sokeri, fosforyloidulla tavalla, muunnetaan entsymaattisesti katalysoiduilla askeleilla varanto- tai rakenteellisiksi polysakkarideiksi.
Lähde: Wesalius [julkinen alue] Triosat osallistuvat lipidien biosynteesiin, jotka ovat osa solukalvoja ja adiposyyttejä.
[TOC]
Ominaisuudet
Glyseraldehydi-aldosassa on quiraalihiiliatomi, ja siksi siinä on kaksi enantiomeroa, L-glyseraldehydi ja D-glyseraldehydi. Molemmilla enantiomeereilla D ja L on erilaiset kemialliset ja fysikaaliset ominaisuudet.
D-glyseraldehydi rikkoo polarisoidun valon tason oikealle (+) ja sillä on kierto [α] D, 25 ° C: ssa, +8: sta.7 °, kun L-glyseraldehydi rikkoutui polarisoidun valon tason vasemmalla (-) ja sillä on kierto [α] D, 25 ° C: ssa, -8: sta.7 °.
Glyseraldehydin kiraalinen hiili on hiili 2 (C-2), joka on toissijainen alkoholi. Fischerin projektio edustaa D-glyseraldehydin hydroksyyliryhmää (-OH) oikealle ja vasemman L-glycen OH-ryhmälle.
Dihydroksiasetonista puuttuu kiraalisia hiiliä, eikä siinä ole enantiomeerisiä muotoja. Hydroksimetyleeniryhmän (-CHOH) lisääminen glyseraldehydiin tai dihydroksiasetoniin mahdollistaa uuden kiraalisen keskuksen luomisen. Tämän seurauksena sokeri on tetrosa, koska siinä on neljä hiilyä.
Ryhmä -chah: n lisääminen Terosaan luo uuden kiraalikeskuksen. Muodostettu sokeri on pentoosi. Ryhmät -CHAH: ta voidaan edelleen lisätä enintään kymmeneen hiilihiiliin.
Voi palvella sinua: fylogenyToiminnot organismissa
Triosat välittäjinä glykolyysissä, glukoneogeneesissä ja pentosa -fosfaattireitissä
Glykolyysi koostuu glukoosimolekyylin repeämästä kahdessa pyruvaattimolekyylissä energian tuottamiseksi. Tämä reitti merkitsee kahta vaihetta: 1) valmisteluvaihe tai energiankulutus; 2) Energiantuotantovaihe. Ensimmäinen on se, joka tuottaa triosan.
Ensimmäisessä vaiheessa glukoosivapaa energiapitoisuus lisääntyy fosfoésterien muodostamalla. Tässä vaiheessa adenosiinitriffosfaatti (ATP) on fosfaatin luovuttaja. Tämä vaihe huipentuu fruktoosifosfoésterin 1,6-bifosfaatin (F1,6 bp) muuntamalla kahdessa triosasfosfaatissa, glyseraldehydi 3-fosfaatissa (GA3P) ja dihydroksiasetonifosfaatissa (DHAP).
Glukoneogeneesi on glukoosibiosynteesi pyruvaatista ja muista välittäjistä. Käytä kaikkia glykolyysentsyymejä, jotka katalysoivat reaktioita, joiden biokemiallinen standardi energian vaihtelu on tasapainossa (ΔGº '~ 0). Tämän vuoksi glykolyysillä ja glukoneogeneesillä on yhteisiä välittäjiä, mukaan lukien GA3P ja DHAP.
Pentoosifosfaattipolku koostuu kahdesta vaiheesta: glukoosi-6-fosfaatin oksidatiivinen faasi ja toinen NADPH- ja Ribosa-5-fosfaatin muodostuminen. Toisessa vaiheessa 5-fosfaatti-riboosi muunnetaan glykolyysivälittäjäiksi, F1.6BP ja GA3P.
Triosas ja Calvin -sykli
Fotosynteesi on jaettu kahteen vaiheeseen. Ensimmäisessä tapahtuu valosta riippuvaisia reaktioita, jotka tuottavat NADPH: n ja ATP: n. Näitä aineita käytetään toisessa, jossa hiilidioksidi ja heksaosin muodostuminen triosasista on kiinnitetty Calvin -syklin kautta.
Calvin-syklissä 1,5-bifhary-oksigenaasi (Rubisco) -ribuloosa-entsyymi katalysoi Co: n kovalenttisen liiton2 Pentoosi ribulosa 1,5-bifosfaatti ja hajottaa kuuden hiiliatomin epävakaan välittäjän kolmen hiiliatomin kahdessa molekyylissä: 3-fosfoglyseraatio.
Voi palvella sinua: Oligosakkaridit: Ominaisuudet, koostumus, toiminnot, tyypitEntsymaattisten reaktioiden kautta, jotka sisältävät 3-fosfoglysereen fosforylaation ja vähentämisen käyttämällä ATP: tä ja NADP: tä, GA3P tapahtuu. Tämä metaboliitti muunnetaan 1,6-bifosfaattfruktoosiksi (F1.6BP) metabolisella reitillä, joka on samanlainen kuin glukoneogeneesi.
Fosfataasin vaikutuksella F1.6BP muunnetaan fruktoosi-6-fosfaatiksi. Sitten isomeasifosfheksoosi tuottaa glukoosin 6-fosfaattia (GLC6P). Lopuksi epicheraasi muuntaa GLC6P: n glukoosi 1-fosfaatiksi, joka palvelee tärkkelyksen biosynteesiä.
Biologisten ja adiposyyttikalvojen triosas ja lipidit
GA3P ja DHAP voivat muodostaa fosfaattiglyserolia, joka on välttämätön metaboliitti trlesyyliglyserolien ja glyserolipidien biosynteesiin. Tämä johtuu siitä, että molemmat triosasfosfaattia voidaan toistaa triosa -isomeraasifosfaatin katalysoimalla reaktiolla, joka pitää molemmat triosat tasapainossa.
Glyserolifosfaattientsyymidehydrogenaasi katalysoi hapettimien vähentämisreaktiota, jossa NADH luovuttaa pari elektronia DHAP: hen muodostaen 3-fosfaattiglyserolin ja NAD: n+. L-glyseroli 3-fosfaatti on osa fosfolipidien luurankoa, jotka ovat biologisten kalvojen rakenteellinen osa.
Glyseroli on proquiraalinen, siitä puuttuu epäsymmetrisiä hiiliä, mutta kun toinen sen kahdesta primaarisesta alkoholista muodostaa fosfoesterin, sitä voidaan oikein kutsua L-glyseroli-3-fosfaatiksi tai D-glyseroli 3-fosfaatiksi.
Glyceofosfolipidejä kutsutaan myös fosfoglyserideiksi, jotka on nimetty fosfatidihapona. Fosfoglyseridit voivat muodostaa fosfalisyyliglyseroleja muodostamalla esterisidoksia kahdella rasvahapolla. Tässä tapauksessa tuloksena oleva tuote on 1,2-fosfodiakyyliglyseroli, joka on tärkeä komponentti kalvoissa.
Glysofosfaasi katalysoi 3-fosfaattiglyserolifosfaattiryhmän hydrolyysiä tuottaen glyserolia enemmän fosfaattia. Glyseroli voi toimia aloitusmetaboliitina trimaktiivisten biosynteesille, jotka ovat yleisiä adiposyyteissä.
Voi palvella sinua: koagulaasitesti: perusta, menettely ja käyttöArkeobakteerien triosas ja kalvot
Samoin kuin eubakteerit ja eukaryootit, triosasfosfaatista muodostuu 3-fosfaattiglyseroli (GA3P ja DHAP). Eroja on kuitenkin: ensimmäinen on, että 3-fosfaattiglyseroli arkeobakteerien kalvoissa on L-konfigurointi, kun taas eubakteereissa ja eukaryoottisissa kalvoissa se on konfiguraatio D.
Toinen ero on, että arkeobakteerikalvot muodostavat esterin yhteydet isoprenoidiryhmien kahteen pitkään hiilivetyketjuun, kun taas eubakteereissa ja eukaryooteissa glyseroli muodostaa esterisidoksia (1,2-diacilglyserolia) kahdella hiilivetyketjulla rasvahappojen ketjuista.
Kolmas ero on, että arkeobakteerikalvoissa fosfaatin ja 3-fosfaatin glyseroli ovat erilaisia kuin eubakteerit ja eukaryootit. Esimerkiksi fosfaattiryhmä on kytketty disakkaridiin α-Glukopiranso- (1®2)-β-Galatofuranosa.
Viitteet
- Cui, s. W -. 2005. Ruokahiilihydraatit: kemia, fysikaaliset ominaisuudet ja sovellukset. CRC Press, Boca Raton.
- Pusku, p., Mätyen, K, Honkala ja., Saag, m., KennePohl, E., Eapen, a. 2016. Erytritoli on tehokkaampi kuin ksylitoli ja sorbitoli suun terveyspäätepisteiden hallinnassa. Kansainvälinen hammaslääketiede.
- Nelson, D. Lens., Cox, m. M. 2017. Lehninger -biokemian periaatteet. W -. H. Freeman, New York.
- Sinnott, m. Lens. 2007. Hiilihydraattikemia ja biokemian rakenne ja mekanismi. Kuninkaallinen kemian yhdistys, Cambridge.
- Rikos. V., Williams, S. J -. 2009. Hiilihydraatit: Elämän olennaiset molekyylit. Elsevier, Amsterdam.
- Voet, D., Voet, J. G., Pratt, c. W -. 2008. Biokemian perusteet - Elämä molekyylitasolla. Wiley, Hoboken.
- « Aldoosiominaisuudet, tyypit, hiilien lukumäärä
- Frooksetiini (PROZAC) vaikutusmekanismi ja sivuvaikutukset »